Advanced pharmaceutical bulletin. 15(3):482-498.
doi: 10.34172/apb.025.43191
Review Article
Genosomes: An Introspection into Transfection, Future Perspectives and Applications
Abhirami Subramony Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing, , # 
Divyatha Raj Data curation, Investigation, Methodology, Writing – original draft, , #
Fiza Fairooz Data curation, Investigation, Methodology, Writing – original draft, , #
Aparna Venugopal Data curation, Investigation, Methodology, Writing – original draft, , #
Aparna Rajesh Data curation, Investigation, Methodology, Visualization, , #
Sreeja C Nair Conceptualization, Supervision, , *
Author information:
Amrita School of Pharmacy, Amrita Institute of Medical Sciences, Amrita Vishwa Vidyapeetham, Kochi-682041, India
#Authors contributed equally to this work.
Abstract
The advent of advanced gene delivery platforms has transformed the precision targeting of therapeutic nucleic acids, such as miRNA, siRNA, and pDNA, for the treatment of genetic and acquired diseases, including cystic fibrosis, malignancies, sickle cell anaemia, and β-thalassemia. Although viral vectors have traditionally dominated this field, non-viral systems, particularly genosomes (cationic lipid-based nanocarriers or lipoplexes), have emerged as promising alternatives due to their enhanced biosafety, lower immunogenic potential, and manufacturability. These nanostructured systems facilitate efficient nucleic acid condensation, protect against enzymatic degradation, and enhance cellular uptake and endosomal escape. Further refinements, including PEGylation, incorporation of helper lipids, and stimuli-responsive formulations, have significantly improved transfection efficiency and tissue-specific delivery. Notable clinical advancements, such as RNA-lipoplexes in cancer immunotherapy and multifunctional envelope-type nanodevices (MEND), highlight their therapeutic potential. This review provides a critical analysis of genosome design strategies, formulation techniques, intracellular trafficking mechanisms, clinical applications, patented innovations, and future prospects to advance genosome-mediated gene therapy.
Keywords: Genosomes, Lipoplexes, Cationic liposome, Transfection efficiency
Copyright and License Information
© 2025 The Author (s).
This is an Open Access article distributed under the terms of the Creative Commons Attribution (CC BY), which permits unrestricted use, distribution, and reproduction in any medium, as long as the original authors and source are cited. No permission is required from the authors or the publishers.
Funding Statement
None.
Introduction
Nucleic acid-based therapeutics have emerged as a transformative modality in modern medicine, offering novel interventions for both inherited and acquired diseases.1,2 These therapies utilise functional genetic materials, such as plasmid DNA (pDNA), small interfering RNA (siRNA), and microRNA (miRNA), to modulate pathological gene expression by restoring defective genes, silencing aberrant transcripts, or reprogramming dysregulated cellular pathways.3 Despite their therapeutic potential, the clinical translation of nucleic acid drugs faces significant delivery challenges.4,5 Unmodified nucleic acids exhibit poor pharmacokinetic profiles due to their high molecular weight, anionic charge, and hydrophilicity, which hinder cellular uptake.6 Additionally, susceptibility to enzymatic degradation and electrostatic repulsion by negatively charged cell membranes further diminishes bioavailability and therapeutic efficacy. While localised administration, such as intramuscular or intratumoral injection, can enhance tissue-specific gene expression, these methods lack the systemic applicability required for many disorders. Consequently, engineered gene delivery vectors have become essential to protect nucleic acids from degradation, enhance cellular internalisation, and enable targeted tissue delivery.
Among the various delivery platforms, lipid-based vesicular systems have gained prominence due to their capacity to encapsulate genetic payloads, improve biodistribution, and facilitate controlled release. Liposomes, nanoscale spherical vesicles composed of lipid bilayers surrounding an aqueous core, represent one of the most extensively studied lipid-based carriers. Their amphiphilic nature allows efficient encapsulation of both hydrophilic and hydrophobic therapeutics, making them versatile vehicles for nucleic acid delivery. Early approaches incorporated viral vectors, including retroviruses and adenoviruses, into liposomal formulations to leverage their high transfection efficiency and nuclear localisation capabilities.7,8 However, despite their initial clinical promise, viral vectors are limited by immunogenicity, insertional mutagenesis (a potential oncogenic risk), toxicity and the challenges associated with large-scale production.9 These limitations have spurred the development of non-viral alternatives, including cationic lipids, polyethyleneimine (PEI)-based polymers, dendrimers, biodegradable polymeric nanoparticles such as PLGA, and cell-penetrating peptides, which offer improved safety, scalability, and tunable physicochemical properties.10
A leading non-viral strategy involves the use of genosomes (or lipoplexes), which are formed through electrostatic complexation between cationic lipids and anionic nucleic acids. These systems present several advantages, including minimal immunogenicity, cost-effective manufacturing, and the ability to preserve the structural and functional integrity of genetic cargo during delivery. Genosomes have demonstrated therapeutic efficacy in treating monogenic disorders (e.g., sickle cell anaemia, β-thalassemia, and cystic fibrosis) as well as various malignancies, including non-small cell lung cancer and breast cancer.11 Current research focuses on optimising genosome formulations through surface modifications, including pegylation and ligand conjugation, and co-delivery with endosomolytic agents to enhance transfection efficiency and tissue specificity.12 With continuous advancements, genosomes represent a clinically viable and versatile platform for next-generation gene therapy, bridging the gap between preclinical innovation and therapeutic application.
Viral vectors v/s non-viral vectors
Viral vectors exhibit unparalleled transfection efficiency, yet their clinical application is hampered by critical safety limitations, including immunogenic responses, cytotoxic effects, and the potential for insertional mutagenesis, a process wherein viral genome integration disrupts tumour suppressor genes or activates proto-oncogenes, increasing the risk of malignant transformation.13,14 In contrast, non-viral delivery systems offer a safer and more controllable alternative, characterised by low immunogenicity, absence of pathogenic risks, cost-efficient manufacturing, and improved biosafety profiles. These advantages, combined with their scalability and long-term stability, have catalysed a shift toward non-viral gene delivery platforms since the early 21st century. A comparative analysis of viral and non-viral vectors is presented in Table 1. While viral vectors, such as adeno-associated viral (AAV), adenovirus (AdV), lentivirus (LV), bacteriophage, and herpes simplex virus (HSV), remain widely utilised in research, non-viral systems are gaining traction in clinical settings. These include lipid-based carriers such as liposomes, cationic polymers (e.g. PEI), inorganic nanoparticles (e.g. gold and silica nanostructures), and endogenous nanovehicles like exosomes and RBC membrane-derived vesicles. Their emerging therapeutic applications are systematically outlined in Tables 2 and 3.
Table 1.
Highlighted differences between viral and non-viral vectors
|
Viral vectors
|
Non-viral vectors
|
| Transfection efficiency is high |
Transfection efficiency is low |
| Causes immunogenicity and cytotoxicity |
It does not cause immunogenicity and cytotoxicity |
| Decreased bio-safety |
Increased bio-safety |
| High cost and difficult production |
Low cost and ease of production |
| Use has gradually decreased due to toxicity |
Use has increased in recent years |
| Not safe to store |
Safe to store |
Table 2.
The most commonly used viral vectors
|
Vectors
|
Family
|
Advantages
|
Disadvantages
|
Ref
|
| Adenoviral |
Adenoviridae |
Titers are large. Can move the transgene into dividing and normal cells |
Transgene expression occurs in a short period and is immunogenic. |
15
|
| AAV |
Parvoviridae |
Abundant host availability, sustainability |
It can produce toxicity, and its packaging capability is restricted. |
16
|
| Retroviral |
Retroviridae |
Safe, low immunogenicity |
Titers are low. Useful for actively dividing cells only. Chances of insertional mutagenesis |
17
|
Table 3.
The most commonly used non-viral vectors
|
Non-viral vectors
|
Example
|
Gene payloads
|
Advantages
|
Applications
|
Ref
|
| Lipid nanoparticles |
Ionisable LNPs (Dlin-MC3-DMA) |
mRNA, siRNA, miRNA, pDNA |
|
Vaccines (e.g. COVID-19), siRNA therapeutics (e.g. Patisiran) |
18
|
| Polymeric nanoparticles |
PEG-PEI, PLGA |
pDNA, CRISPR-Cas9, shRNA |
|
Cancer gene therapy, regenerative medicine |
19
|
| Inorganic nanoparticles |
Gold nanoparticles, mesoporous silica |
siRNA, ASO, mRNA |
|
Targeted therapy like tumour-specific delivery |
20
|
| Cell-penetrating peptides |
TAT peptide, Penetratin |
CRISPR RNP, siRNA, pDNA |
|
Neurological disorders, genome editing |
21
|
| Hydrogels/ Hybrid systems |
Chitosan-hyaluronic acid |
mRNA, pDNA, miRNA |
|
Tissue engineering, localised therapy |
22
|
Nucleic acids utilised in Genosome-based delivery systems
The selection and optimisation of nucleic acid payloads represent a critical determinant in the efficacy of genosome-based gene delivery systems, with each class of therapeutic oligonucleotides, including siRNA, miRNA, ASOs, and CRISPR-Cas9 components, presenting unique physicochemical properties and delivery challenges that necessitate tailored formulation strategies (Table 4).
Table 4.
Comparative analysis of nucleic acids for delivery by genosomes
|
Category
|
siRNA
|
miRNA
|
ASOs
|
CRISPR-Cas9 system
|
| Size (nt/kb) |
20-25 |
20-22 |
15-25 |
gRNA: ~100
Cas9: ~4000 |
| Charge |
Strongly negative |
Negative |
Modifiable |
Negative (RNA)
Variable (Protein) |
| Key delivery challenge |
Nuclease sensitivity
Endosomal escape |
Tissue specificity
Off-target effects |
Nuclear delivery
Biodistribution |
Payload size
Immunogenicity |
| Genosome as a delivery vehicle |
Ionisable LNPs
Cationic complexation |
Targeted formulations
PEGylation |
Stabilised formulations
Nuclear localisation signals |
Co-delivery systems
pH-sensitive lipids |
| Clinical relevance |
FDA-approved formulations, e.g. Patisiran |
MRX34 |
FDA-approved formulations, e.g. Nusinersen |
NTLA-2001 |
| Therapeutic advantage |
Potent silencing
Reversible effect |
Multi-gene modulation
Natural regulator |
Splice modulation
Chemical versatility |
Permanent editing
Precision targeting |
| Optimal lipid composition |
Dlin-MC3-DMA
DSPC: Chol: PEG |
DOTAP: DOPE
Targeted PEG-lipids |
Neutral/stealth lipids
Phosphorothioate analogs |
Cationic: neutral blends
Helper lipids |
| Reference |
23
|
24
|
25
|
26
|
Small interfering RNA (siRNA)
siRNA has become a prime candidate for genosome-mediated delivery owing to its well-defined RNA interference (RNAi) mechanism and compact molecular structure. The 20-25 base pair duplexes readily form stable complexes with cationic lipids via electrostatic interactions, enabling efficient encapsulation within lipid nanoparticles (LNPs).27 Genosomes overcome major siRNA delivery barriers by providing nuclease protection during systemic circulation and enhancing cellular uptake through charge-mediated endocytosis.28,29 Advanced formulations employ ionisable lipids (e.g. Dlin-MC3-DMA) that undergo pH-dependent protonation in endosomes, facilitating membrane destabilisation and cytosolic siRNA release. This strategy has achieved clinical validation with Patisiran (Onpattro®), an FDA-approved genosome, delivered siRNA therapeutic for transthyretin-mediated amyloidosis that demonstrates effective hepatocyte-specific gene silencing.30
MicroRNA (miRNA)
As endogenous post-transcriptional regulators, miRNA presents both therapeutic opportunities and delivery challenges for genosome systems.31 While their ability to modulate multiple disease pathways simultaneously is advantageous, the risk of off-target effects necessitates precise tissue targeting.32 Contemporary genosome designs address these requirements through surface-conjugated targeting ligands (e.g. folate, RGD peptides) for cell-specific delivery, and PEGylation to enhance circulatory half-life by minimising reticuloendothelial clearance. The amphiphilic nature of genosomes permits stable incorporation of miRNA mimics or antagomirs while preserving biological activity.33 Current optimisation efforts focus on lipid composition refinements to improve tissue accumulation profiles, as exemplified by clinical-stage candidates like MRX34 for oncology applications.
Antisense oligonucleotide (ASO)
Genosome encapsulation significantly enhances the therapeutic potential of ASOs by addressing their delivery limitations.34 Although chemical modifications like phosphorothioate backbones, 2’-O-methyl groups improve ASO stability and target affinity, they remain insufficient for efficient intracellular delivery.35 Genosome formulations provide comprehensive solutions by protecting ASOs from serum nucleases, promoting cellular internalisation through optimised surface charge, and facilitating nuclear localisation via incorporated targeting motifs.36,37 While ASO therapies like Nusinersen (Spiranza®) for spinal muscular atrophy have demonstrated clinical success, genosome delivery could further enhance tissue distribution and pharmacokinetics, potentially reducing dosing frequency.38
CRISPR/Cas9
The CRISPR/Cas9 platform presents unique delivery challenges that genosomes are particularly equipped to address.39 Unlike smaller nucleic acids, CRISPR components require co-delivery of both guide RNA (~100 nt) and Cas9 protein/mRNA (~4 kb coding sequence), demanding substantial payload capacity. Genosomes meet these requirements through cationic lipid-mediated nucleic acid condensation, ionisable lipid-facilitated endosomal escape, and optimised lipid ratios for particle stability.40 While clinical trials have demonstrated successful hepatic delivery, achieving efficient extrahepatic targeting remains challenging.41 Current research focuses on improving tissue specificity and reducing immune recognition.
Structure of a genosome
Genosomes represent a class of synthetic, lipid-based nanocomplexes engineered for nucleic acid delivery, comprising three essential components- a cationic lipid, a neutral helper lipid, and the therapeutic payload (DNA or RNA).42,43 These nanostructures derive their stability from electrostatic interactions between the positively charged lipid moieties and the anionic phosphate backbone of the nucleic acids, thereby ensuring both structural integrity and protection of the genetic cargo.44,45 The cationic lipoplex system, a fundamental architectural element of genosomes, enhances transfection efficiency through charge-mediated interactions.46 The positively charged nanoparticle surface facilitates cellular uptake via association with negatively charged cell surface proteoglycans. Following internalisation, controlled disassembly of the complex enables targeted release of the nucleic acid payload, either to the cytoplasm (for RNA-based therapeutics) or the nucleus (for DNA-based therapeutics), to mediate gene silencing or expression. At the molecular level, genosomes adopt a spherical bilayer morphology, wherein amphiphilic lipids self-assemble with their hydrophobic tails oriented outward and hydrophilic headgroups inward (Figure 1). The cationic character of these lipids, typically conferred by primary or quaternary ammonium groups, serves dual functions- electrostatic condensation of nucleic acids, and promotion of cellular uptake.47,48 Notably, the number and chemical nature of these ammonium groups critically influence both nucleic acid compaction efficiency and overall transfection performance.
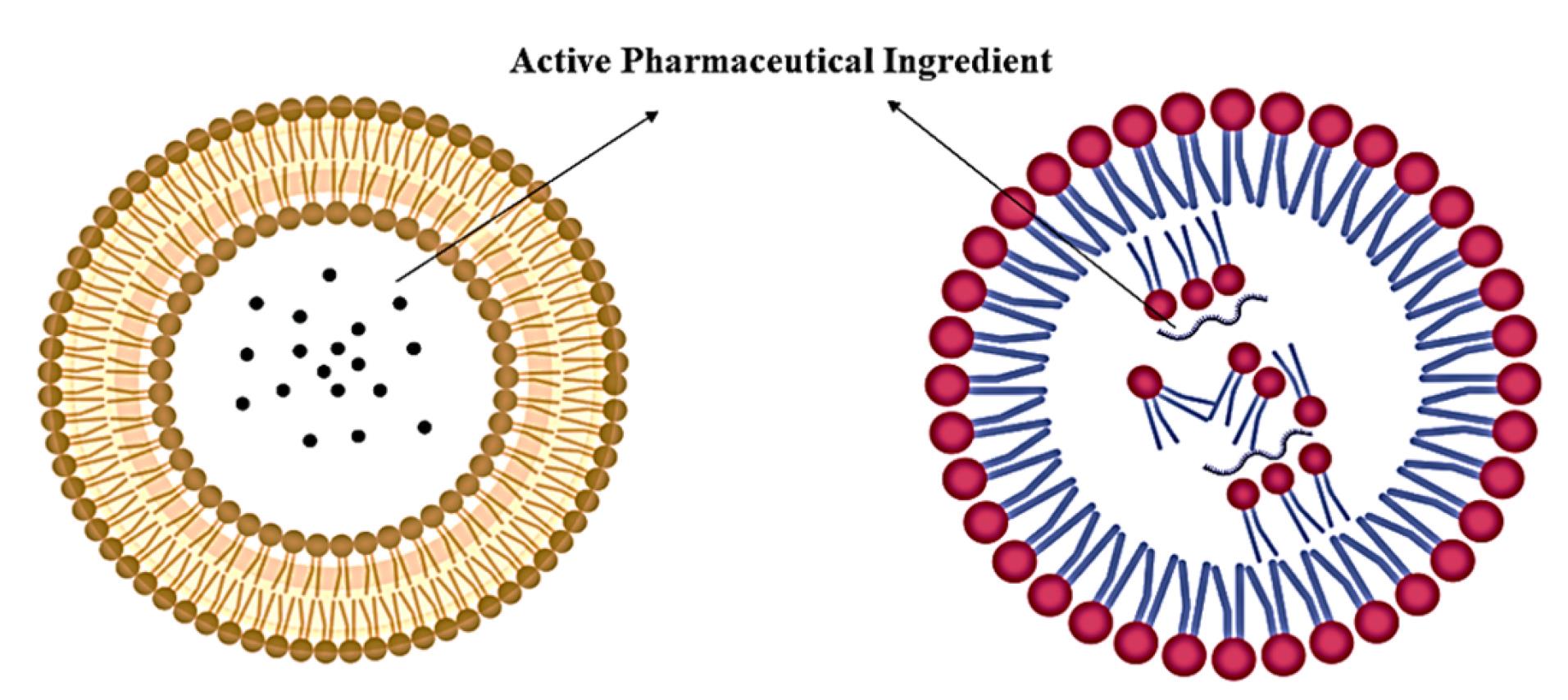
Figure 1.
Structures of liposome and lipid nanoparticle
.
Structures of liposome and lipid nanoparticle
Genosomes exhibit dynamic phase behaviours, predominantly adopting lamellar (Lα) or inverted hexagonal (H_II) arrangements at the nanoscale (10-50 nm), irrespective of the preparation method. In the lamellar phase, nucleic acids intercalated between parallel lipid bilayers in a rod-like conformation, with membrane fluidity modulated by temperature and lipid composition.49 H_II phase organises nucleic acids within a two-dimensional hexagonal lattice, while the micellar hexagonal phase localises them in interstitial, honeycomb-like regions.50,51 More complex architectures, including cubic and bicontinous phases, have also been documented.
At larger length scales (100 nm to 1 μm), genosome morphology is dictated by preparation parameters such as mixing order and assembly conditions.52,53 While nanoscale organisation governs transfection efficiency, macroscale features influence critical in vivo performance metrics, including circulation half-life, biodistribution, and plasma stability.54,55
The structural organisation of genosomes is primarily described by two theoretical models.56,57 The external binding model proposes that nucleic acids adsorb onto the lipid nanoparticle surface, forming a distinctive “beads-on-a-string” morphology stabilised by electrostatic interactions between the cationic lipids and anionic nucleic acids.58,59 In contrast, the internal encapsulation model suggests complete entrapment of nucleic acids within the lipid core, resulting in multilamellar or vesicular architectures that provide enhanced protection to the genetic payload.60 Beyond these primary configurations, specialised lipid formulations like SAINT-2 lipids can generate alternative structural variants such as ellipsoidal genosomes, which form through rapid, thermodynamically favourable self-assembly processes when cationic vesicles complex with pDNA.61 These diverse structural paradigms collectively influence the stability, release kinetics and transfection efficacy of genosome-based delivery systems.
Formation of genosomes
Genosomes are synthesised through electrostatic complexation between nucleic acids and cationic lipid formulations, as illustrated in Figure 2.62 The assembly process involves mixing lipid and nucleic acid components in an aqueous medium under ambient conditions (20–25 °C), where the positively charged amine groups of cationic lipids interact with the anionic phosphate backbone of nucleic acids, a critical determinant of transfection efficiency.63 Empirical studies confirm that higher surface charge density correlates with enhanced transfection capacity due to improved nucleic acid binding affinity.64
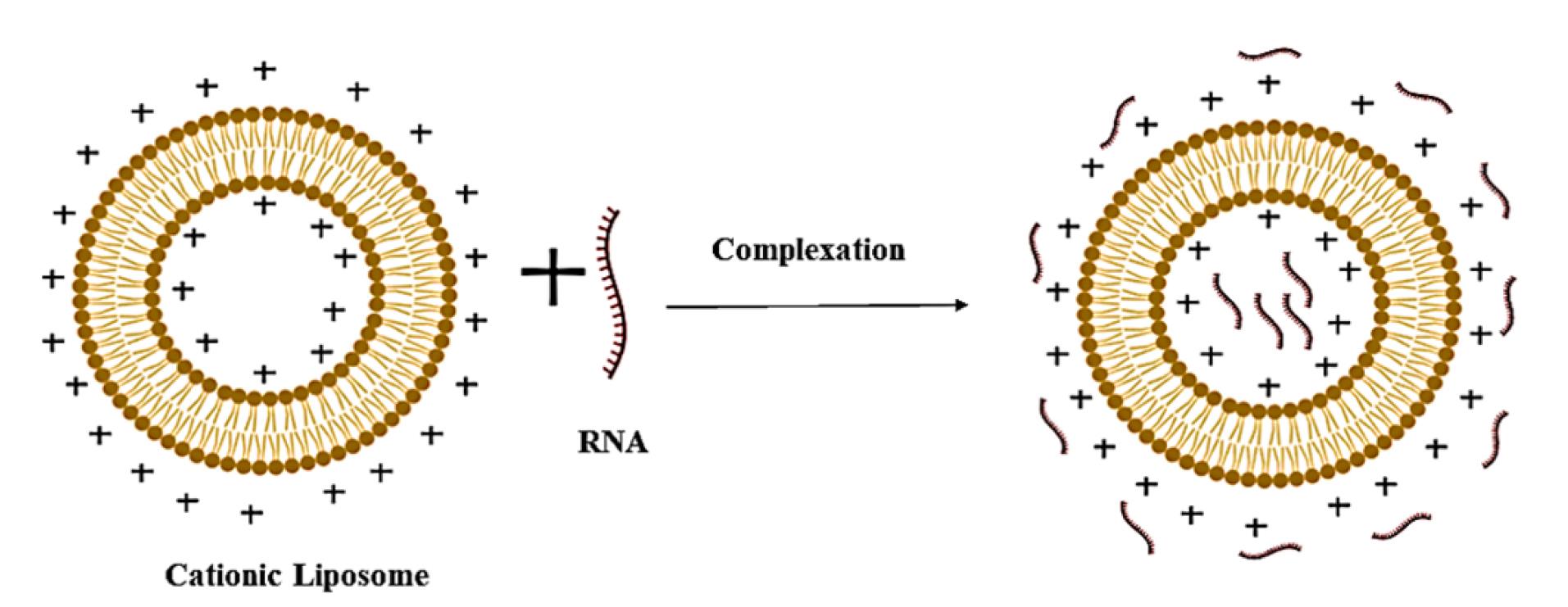
Figure 2.
Schematic representation of genosome formation
.
Schematic representation of genosome formation
The formation mechanism occurs via two distinct thermodynamic phases.65 The initial step involves rapid ( < 1 h), exothermic adsorption of nucleic acids onto cationic lipid surfaces, accompanied by counterion release (~90% from lipids and ~70% from nucleic acids), which stabilises the complex through electrostatic bridging.66,67 Subsequently, a slower, endothermic reorganisation phase mediates nucleic acid encapsulation within the lipid matrix. This irreversible process entails disruption of hydrophobic lipid domains followed by structural stabilisation via van der Waals forces, ultimately defining the lipoplex architecture and biological performance.68,69
Commonly employed cationic lipids, including DOTMA, DOTAP, DOGS, DODAC, and DODMA, are selected based on their charge density and fusogenicity, while helper lipids (e.g. DOPE, cholesterol (CHOL) enhance membrane stability and cellular uptake.70,71 As summarised in Table 5, these lipids exhibit distinct performance profiles. For instance, DOTAP demonstrates moderate transfection efficiency (65±10%) but significant cytotoxicity, whereas ionisable lipids like Dlin-MC3 achieve superior efficiency (80±5%) with reduced toxicity. Particle size, governed by the lipid: nucleic acid charge ratio, critically influences functionality. Near-neutral ratios (slight cationic excess) generate larger complexes with enhanced transfection efficiency, while extreme charge ratios produce smaller but less effective particles.72 Notably, excess free liposomes must be removed post-formulation to mitigate cytotoxicity, ensuring optimal safety and therapeutic efficacy.
Table 5.
Comparative performance of genosome formulations
|
Lipid type
|
Particle size
|
Zeta potential
|
Uptake (vs control)
|
Example of formulation
|
Clinical relevance
|
Advantages
|
Limitations
|
Ref.
|
| Cationic (e.g. DOTAP/DOPE) |
~100-150 nm |
Highly positive (~ + 30 mV) |
3-4x higher |
siRNA for lung cancer |
Localised therapy (e.g. intratumoral) |
Rapid cell entry
high DNA loading |
Toxic at high doses
Unstable in the blood |
73
|
| Ionisable (e.g. Dlin-MC3) |
~80-100 nm |
pH-sensitive (neutral in blood, positive in endosomes) |
4-5x higher |
Onpattro® (hATTR amyloidosis) |
Systemic delivery (liver-targeted) |
FDA-approved formulations
Low immune reaction |
Requires cold chain storage
Expensive production |
74
|
| PEGylated Lipids |
~150-200 nm |
Slightly positive (~ + 10 mV) |
2-3x higher |
mRNA vaccines (COVID-19 LNPs) |
Vaccines/ repeat dosing |
Long blood circulation
Stealth effect |
Adverse effects like PEG allergy in some patients |
75
|
| Neutral (e.g. DOPE/Chol) |
~180-250 nm |
Near-neutral |
1.5-2x higher |
Gene therapy for brain diseases |
Neurodegenerative disorders |
Very low toxicity
Biocompatible |
Poor gene release from endosomes |
76
|
| Targeted (e.g. Folate-PEG) |
~90-120 nm |
Slightly negative (~ -5mV) |
5-6x higher (in cancer cells) |
Ovarian cancer therapy |
Receptor-positive cancers |
Tumour-selective
Minimal side effects |
Limited to receptor-specific
targeting |
77
|
Intracellular delivery mechanisms of genosome-based therapeutics
The therapeutic efficacy of genosome-mediated gene delivery systems fundamentally depends on their capacity to achieve successful intracellular trafficking and payload release within target cells.78 This complex biological process occurs through two mechanistically coupled phases- receptor-mediated cellular internalisation, followed by endosomal escape.79 Cellular uptake is initiated when cationic lipoplexes interact with negatively charged cell surface proteoglycans, primarily through clathrin-mediated endocytosis (CME), although alternative pathways, including caveolae-dependent uptake and micropinocytosis, may contribute to varying degrees depending on the specific formulation characteristics. These endocytic mechanisms not only protect the nucleic acid payload from extracellular nucleases but also facilitate transport into the acidic environment of endolysosomal compartments (pH 5.0-6.5), where critical structural transformations occur.80,81
The endosomal escape process presents the most formidable barrier to successful gene delivery, with two well-characterised mechanisms currently proposed.82,83 The flip-flop model involves a sophisticated lipid exchange process where anionic phospholipids (particularly phosphatidylserine) from the inner endosomal membrane leaflet translocate to the outer leaflet through ATP-dependent flippase activity (Figure 3). This translocation creates charge-neutralised ion pairs between the anionic phospholipids and cationic lipids of the genosome, significantly reducing the electrostatic binding affinity for nucleic acids. The resulting charge neutralisation induces a phase separation in the lipoplex structure, ultimately leading to nucleic acid release into the cytosol.84 However, this mechanism demonstrates limited efficiency due to kinetic constraints in lipid exchange rates and geometric mismatches between the genosome surface area and available endosomal membrane phospholipids.85
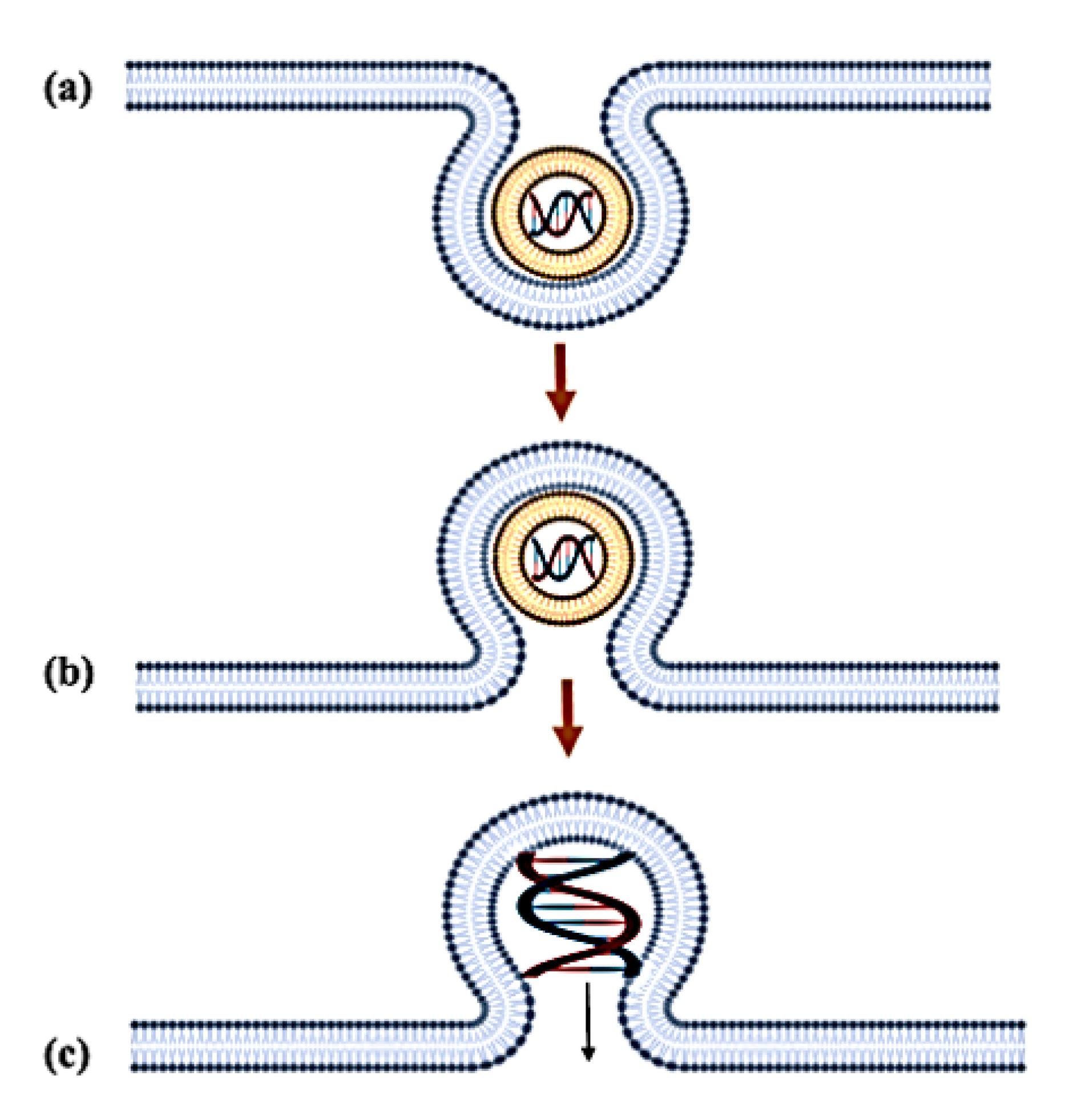
Figure 3.
Flip-flop mechanism of the genosome: (a) Fusion of lipoplex with the endosomal membrane (b) Flip-flop of the negatively charged phospholipids (c) Destabilisation of the membrane and release of the nucleic acid
.
Flip-flop mechanism of the genosome: (a) Fusion of lipoplex with the endosomal membrane (b) Flip-flop of the negatively charged phospholipids (c) Destabilisation of the membrane and release of the nucleic acid
More recent studies have provided substantial evidence for the transient pore formation mechanism, which involves a multi-step process of membrane destabilisation (Figure 4).86 Initially, genosomes localise to the inner endosomal membrane through electrostatic interactions, followed by partial degradation of the lipid-nucleic acid complex in the acidic environment. This degradation induces localised membrane curvature stress, leading to the formation of transient hydrophilic pores (3-10 nm diameters) with lifetimes ranging from milliseconds to seconds. Molecular dynamics simulation suggests these pores are stabilised by the inverted cone-shaped geometry of the helper lipids like DOPE, which reduces the energetic barrier for pore formation. The nucleic acid payload escapes through these transient defects via a combination of electrophoretic forces and concentration gradients, after which the membrane rapidly reseals through lipid rearrangement. This mechanism appears particularly efficient for larger nucleic acid payloads such as pDNA and CRISPR-Cas9 complexes, potentially explaining the superior transfection efficiency observed with certain lipid formulations.
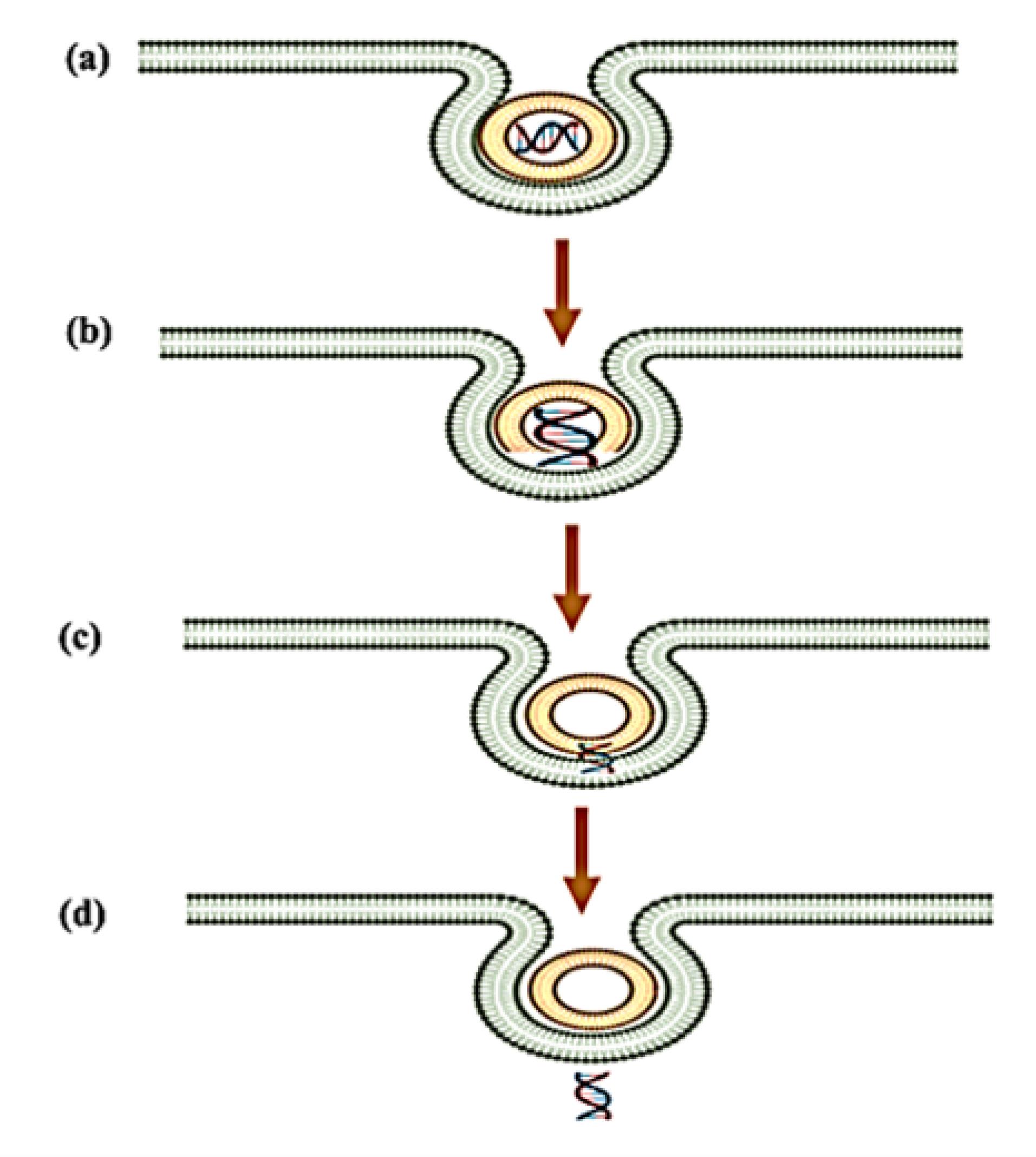
Figure 4.
Transient pore formation: (a) Genosome gets localised to the inner-endosomal membrane (b) Dispersal of lipid molecule by lipoplex degradation (c) Endosomal migration of the nucleic acid by transient pore formation (d) Slow release of the nucleic acid after lipid integration
.
Transient pore formation: (a) Genosome gets localised to the inner-endosomal membrane (b) Dispersal of lipid molecule by lipoplex degradation (c) Endosomal migration of the nucleic acid by transient pore formation (d) Slow release of the nucleic acid after lipid integration
These sophisticated delivery mechanisms ensure appropriate subcellular localisation of therapeutic nucleic acids while minimising lysosomal degradation. Current optimisation strategies focus on engineering lipid compositions with improved endosomolytic activity, including the development of pH-sensitive ionisable lipids and the incorporation of endosomolytic peptides, while maintaining favourable safety profiles for clinical applications.
In vitro/in vivo comparison of endosomal escape mechanisms in genosome delivery
The flip-flop and transient pore mechanisms represent two distinct but potentially complementary pathways for genosome-mediated endosomal escape, each with unique biophysical characteristics and functional implications. In physiological systems, the flip-flop mechanism dominates, accounting for approximately 60% of release events, as demonstrated by live-imaging studies in primate models.87 This process involves the ATP-dependent reorganisation of membrane phospholipids where anionic species translocate across the bilayer to neutralise cationic lipids, ultimately facilitating nucleic acid release. The biological relevance of this mechanism is underscored by its dependence on endogenous lipid recycling pathways that are characteristic of living systems. In contrast, in vitro environments favour pore formation, where static culture conditions promote the generation of transient 5-10 nm membrane defects that enable cytosolic entry, as visualised through cryo-EM studies. This pore-mediated escape demonstrates higher efficiency (80% in HeLa cells) but may overestimate delivery potential due to the absence of physiological barriers present in living organisms.88
At the molecular level, these mechanisms exhibit distinct characteristics with important clinical implications. The flip-flop process involves complex lipid rearrangements and salt bridge formation, with FRET-based assays confirming nanoscale reorganisation events. This mechanism’s physiological fidelity makes it particularly valuable for therapeutic development, as evidenced by the superior performance (2-fold higher gene expression) of flip-flop-optimised LNPs in clinical settings.89 Meanwhile, pore formation occurs through passive membrane strain, generating short-lived ( < 1 second) leakage pathways detectable by advanced imaging techniques like atomic force microscopy.90 Current research reveals these mechanisms may not be mutually exclusive, with emerging hybrid models suggesting pore formation may initiate flip-flop by exposing inner membrane leaflets. Computational studies further indicate potential lipid-protein cooperatively in pore stabilisation, adding complexity to our understanding of these processes.91
Significant questions remain regarding precise molecular requirements and the dynamic interplay of these escape mechanisms in living systems. Key challenges include determining the exact lipid stoichiometry necessary for efficient flip-flop in vivo and developing methods to quantitatively track pore dynamics in physiological environments. Recent advances in multiplexed imaging and AI-assisted molecular dynamics simulations are beginning to address these knowledge gaps. These investigations are crucial for optimising next-generation genosome designs, particularly in achieving the delicate balance between efficient endosomal escape and minimal cytotoxicity, a critical factor in translating nanocarrier systems from bench to bedside. The continued elucidation of these escape mechanisms will undoubtedly enhance our ability to engineer more effective and targeted gene delivery systems for diverse therapeutic applications.
Functional role of helper lipids in genosome systems
Helper lipids serve as critical structural and functional components of genosome formulations, with DOPE and DOPC being most widely utilised for their biomimetic properties and pH-responsive behaviour.92,93 DOPE’s ability to undergo lamellar-to-hexagonal (Lα-to-H_II) phase transition under endosomal acidic conditions (pH 5.0-6.5) promotes membrane destabilisation through curvature stress induction and non-bilayer intermediate formation. Molecular interactions between phosphate groups of DOPE and cationic lipid headgroups enhance nucleic acid condensation while facilitating endosomal escape, with optimal activity observed at 30-50 mol% concentrations. Cholesterol further stabilises the lipid bilayer through hydrophobic interactions and modulates membrane fluidity for improved systemic circulation.94
Recent advances have transformed helper lipids from passive structural elements to active functional components through engineered features like pH-triggered conformational switches and intracellular trafficking motifs. These developments enable precise control over genosome stability, biodistribution, and intracellular release kinetics. Current research focuses on synthetic analogues with tunable phase behaviour and reduced immunogenicity, representing a key strategy for optimising the therapeutic index of genosome-based delivery systems while addressing critical translational challenges. The rational design of helper lipid compositions now stands as a fundamental parameter in developing clinically viable nucleic acid therapeutics.
Advanced lipid design strategies for enhanced genosome performance
Recent innovations in lipid engineering have yielded significant improvements in genosome-mediated transfection efficiency through rational molecular design. A particularly impactful approach involves the development of pH-responsive cationic lipids containing ionisable amine or imidazole groups.95 These smart lipids exhibit precisely tuned pKa values (6.2-6.6) that enable charge-state transitions in response to endosomal acidification.96 The pH-dependent protonation of these functional groups induces two critical effects- increases electrostatic interactions with endosomal membranes, and structural transitions that promote membrane destabilisation, collectively enhancing nucleic acid release into the cytosol.
Further optimisation has been achieved through strategic incorporation of unsaturated fatty acid chains (e.g. oleic and linoleic acids) within the lipid architecture.97 The cis-double bonds in these hydrocarbon tails introduce kinks that reduce packing density and increase bilayer fluidity. These enhanced membrane dynamics facilitates three key processes- improved fusion with cellular membranes, more efficient disassembly of lipid-nucleic acid complexes, and optimised intracellular trafficking of genetic payloads. Current research focuses on combinatorial approaches that integrate these design principles with targeting ligands and stealth components to develop next-generation genosomes with superior tissue specificity and transfection efficiency.
Surface engineering through PEGylation
PEGylation, the covalent conjugation of polyethylene glycol (PEG) to genosome surfaces, represents a critical strategy for enhancing systemic circulation and reducing immune clearance.98 The hydrophilic PEG chains form a static barrier that minimises opsonisation and recognition by the mononuclear phagocyte system, thereby extending plasma half-life. However, this shielding effect presents a dual challenge- excessive PEGylation can impair critical interactions with cellular membranes, reducing endocytic uptake and intracellular delivery efficiency.99
To address these limitations, advanced PEGylation strategies employ short-chain unsaturated PEG derivatives that balance stealth properties with subsequent detachment in the target microenvironment. These optimised formulations maintain sufficient PEG density for initial immune evasion while allowing timely PEG shedding to facilitate membrane fusion and cellular internalisation. Although this approach may modestly reduce circulation time compared to conventional PEGylation, it significantly enhances transfection efficiency by preserving the genosome’s ability to interact with target cells. Current research focuses on stimuli—responsive PEG lipid conjugates that undergo controlled deshielding in response to tumour microenvironment cues (e.g. pH, enzymes) for improved spatiotemporal delivery control.
Physiological barriers in genosome delivery
The efficacy of genosome-based gene delivery is governed by their ability to overcome multiple physiological barriers encountered during systemic administration.100 Extracellular challenges include enzymatic degradation, serum protein adsorption, and immune surveillance mechanisms that can rapidly clear nanoparticles from circulation. Upon cellular internalisation, genosomes must subsequently navigate a series of intracellular obstacles- plasma membrane penetration, endosomal escape, cytoplasmic trafficking, and, for DNA-based therapeutics, nuclear entry, all while maintaining payload integrity.101 A critical additional requirement involves controlled unpacking of the nucleic acid cargo to ensure proper temporal release for optimal gene expression or silencing activity.
To address these challenges, surface engineering strategies have been developed to improve genosome pharmacokinetics and biodistribution. Charge-shielding modifications using neutral or hydrophilic polymers such as galactose, dextran, or PEG effectively reduce non-specific interactions with plasma proteins, particularly albumin, and minimise immune recognition. These modifications must be carefully balanced to maintain sufficient cellular interaction while preventing rapid clearance. Current optimisation approaches integrate stimuli-responsive elements that maintain stealth properties during circulation but undergo controlled activation at target sites. The successful clinical translation of genosome technology ultimately depends on this multifaceted design paradigm that simultaneously addresses lipid composition optimisation, controlled release mechanism, and physiological barrier evasion, all critical factors for achieving targeted, safe, and effective gene therapy outcomes.
Parameters influencing physico-chemical properties of genosomes
The therapeutic performance of genosome systems is fundamentally governed by their transfection efficiency, which is directly modulated by key physicochemical properties including surface charge density, particle size distribution, and colloidal stability.102 These characteristics emerge from complex thermodynamic and kinetic interactions during formulation, where critical parameters such as lipid-to-nucleic acid ratio, ionic strength of the medium, mixing kinetics, and complexation temperature must be precisely controlled. The lipid-to-DNA charge ratio represents a particularly crucial thermodynamic parameter that dictates the structural organisation of resulting lipoplexes- highly cationic complexes achieve complete nucleic acid condensation, while anionic formulations often contain unbound DNA strands.103 Neutral charge ratios (zeta potential ~ 0) typically produce heterogeneous size distributions due to diminished interparticle electrostatic repulsion, leading to aggregation and compromised stability.
Advanced empirical approaches have been developed to control genosome characteristics through careful manipulation of preparation conditions.104 Charge-directed assembly methods demonstrate that adding nucleic acids to preformed lipid dispersions yields positively charged complexes, while inverse addition produces anionic systems. Mixing kinetics significantly influence particle morphology, with rapid mixing generating smaller, more uniform lipoplexes compared to the aggregated structures formed during slow mixing. While temperature effects are generally minimal for DNA stability, they may modulate complexation kinetics. Ionic strength represents another critical variable, where elevated salt concentrations can both promote component association through charge screening while potentially destabilising colloids. These formulation parameters must be systematically optimised to achieve the delicate balance between nucleic acid protection, cellular uptake efficiency, and intracellular release kinetics required for effective lipofection. Current research focuses on computational modelling approaches to predict optimal formulation conditions based on molecular interaction parameters, representing a promising direction for rational genosome design.
Application of genosomes
Genosome-based therapeutics have achieved notable clinical success since their first regulatory approval in 2003, when China approved Gendicine for the treatment of head and neck squamous cell carcinoma.105,106 These lipid-based non-viral vectors represent a paradigm shift in gene therapy, particularly for genetic disorders and refractory cancers where conventional treatments prove inadequate. Their design leverages cationic lipids to condense nucleic acids while mimicking viral delivery mechanisms, offering distinct advantages including reduced immunogenicity, lower toxicity profiles, and improved targeting capabilities compared to viral vectors- albeit with generally lower transfection efficiency.107 The clinical development of genosomes has progressed significantly across multiple therapeutic areas through innovative formulation strategies.
In pulmonary medicine, genosomes enable cell-specific delivery through optimised administration routes. Intranasal delivery of SPC-targeted miRNA lipoplexes achieves selective transfection of alveolar type II pneumocytes, demonstrating enhanced local action at the airway epithelium.108 The multifunctional envelope-type nano-device (MEND) represents a significant advancement, with the optimised YS05-MEND formulation showing superior efficacy against lung metastases compared to conventional chemotherapy in preclinical models.109,110 These systems combine lipid, polymer, and protein components to enhance cytosolic delivery of RNA therapeutics while maintaining favourable safety profiles.
Genosome technology shows particular promise for cutaneous applications where localised delivery is paramount. For radiation-induced skin damage, PUMA siRNA incorporated into Carbopol hydrogels enables epidermal-specific RNAi delivery while sparing deeper tissue layers.111 Co-formulation with DOTAP further enhances transdermal delivery to epidermal melanocytes, demonstrating the importance of lipid composition in tissue penetration.112 Hair follicle gene therapy represents another innovative application, with in vivo murine studies achieving up to 50% transfection efficiency in progenitor cells following depilation and retinoic acid pretreatment.113 These applications highlight the critical relationship between formulation parameters such as lipid-to-DNA ratio and absolute concentrations and therapeutic outcomes.114
Genosomes address the formidable challenge of blood-brain barrier (BBB) penetration for neurological applications. Lipoplex systems combining lipid and polymer components enable non-invasive delivery of therapeutic genes to the CNS while maintaining high loading capacity and production scalability.115,116 Advanced targeting strategies include transferrin receptor-targeted immunoliposomes, which in recent studies restored striatal tyrosine hydroxylase activity in Parkinson’s disease models.117,118 For Alzheimer’s disease (AD), chitosan-precondensed lipoplexes delivering ApoE2 plasmids show therapeutic potential, with bifunctionalised systems (mApoEPA-LIP) demonstrating reduced amyloid burden and cognitive improvement in transgenic models.119,120 Surface modification targeting GLUT1 transporters and incorporating rabies virus glycoprotein derivative further enhances CNS penetration.121
Genosome formulations overcome significant pharmacological challenges in treating parasitic infections. Dinitroaniline compounds, while effective against Leishmania species, benefit from PC-based encapsulation to address poor solubility and stability issues.122 In malaria therapeutics, soy-PC and cholesterol lipoplexes containing monensin demonstrate enhanced activity against both Plasmodium berghei in murine models and Plasmodium falciparum in vitro.123,124 These applications underscore the importance of lipid composition of genosomes in improving drug bioavailability and therapeutic index for infectious diseases
RNA-lipoplexes represent a breakthrough in cancer immunotherapy, with several candidates reaching clinical trials. These formulations specifically target splenic dendritic cells to enhance antigen presentation, as demonstrated in B16 melanoma models where tumour progression was significantly suppressed.125,126 The structural transition from RNA-cationic liposome complexes to RNA-lipoplexes provides critical protection against nucleases while improving cellular uptake. Vascular-targeted siRNA lipoplexes show similarly promising results, addressing the rapid clearance limitations of free siRNA through enhanced tissue retention and favourable pharmacokinetics without observed toxicity.
Advances in genosome therapeutics: Clinical progress, delivery challenges and patent landscape
Genosome-based therapies have revolutionised precision medicine. With landmark approvals such as Patisiran (Onpattro®) for hereditary transerythrin amyloidosis (hATTR) and mRNA-LNP vaccines for COVID-19, these platforms now target oncology, genetic disorders, and infectious diseases. However, challenges in delivery efficiency, immunogenicity, and manufacturing scalability persist.
Clinical progress in genosome therapeutics
Recent years have witnessed transformative clinical advancements in genosome-based therapeutics, particularly in oncology, genetic disorders, and infectious diseases. In oncology, siRNA and mRNA platforms have demonstrated promising antitumour activity, albeit with challenges in tolerability. For instance, a Phase I trial investigating LNP-encapsulated KRAS-targeting siRNA in pancreatic cancer reported significant tumour regression in 30% of patients, although dose-limiting cytokine release syndromes necessitated careful dose optimisation.127 Similarly, personalised neoantigen mRNA vaccines have shown enhanced T-cell responses in melanoma, underscoring the potential of mRNA-LNPs in cancer immunotherapy. Beyond oncology, CRISPR-based therapies for hereditary transthyretin amyloidosis achieved substantial serum TTR reduction, yet liver toxicity remains a critical barrier to broader application.128
The clinical development of genosome therapies remains highly skewed toward early-phase trials, with over 80% of ongoing studies in Phase I/II. This trend reflects the emphasis on safety assessments, particularly for novel modalities like CRISPR and tumour-targeted siRNA. However, notable late-stage successes, such as Patisiran’s approval for hATTR and the rapid deployment of mRNA-LNP COVID-19 vaccines, validate the translational potential of these platforms.129 The APOLLO trial established Patisiran’s 0.3 mg/kg every-three-weeks regimen as optimal, achieving an 81% reduction in pathogenic TTR levels, while the COVID-19 vaccines highlighted the pivotal role of ionisable lipids (e.g. SM-102, ALC-0315) in enhancing efficacy and stability.130
Despite these successes, key challenges persist, including cytokine-driven toxicities in oncology applications, immunogenicity of PEGylated lipids, and limited extrahepatic delivery efficiency.131 Table 6 summarises recent landmark clinical trials, illustrating the therapeutic scope and unresolved hurdles in genosome development. Addressing these limitations through innovative delivery strategies and robust safety monitoring will be critical for advancing next-generation candidates into late-phase trials and clinical practice.
Table 6.
Current status of genosome-based therapies in clinical trials
|
Delivery system
|
Gene
|
Delivery route
|
Indications
|
Development phase
|
Status
|
Sponsor
|
ClinicalTrials.gov identifier
|
Ref.
|
| Pbi-shRNA lipoplex |
EWS/FLI1 gene (driver gene of Ewing’s sarcoma) |
Intravenous |
Advanced Ewing’s sarcoma |
1 |
Active, not recruiting |
Gradalis, Inc. |
NCT02736565 (2023) |
132
|
| Pbi-shRNA lipoplex |
STMN 1 gene (leukaemia-associated cytoplasmatic phosphoprotein) |
Intratumoral |
Advanced and/or Metastatic cancer |
1 |
Completed |
Gradalis, Inc. |
NCT01505153 (2018) |
133
|
| Tetravalent RNA-lipoplex |
NY-ESO-1 (New York ESO-1) MAGE-A3 (Melanoma associated antigen A3) TPTE (trans-membrane phosphatase with tensin homology) Tyrosinase |
Intravenous |
Advanced Melanoma |
1 |
Active, not recruiting |
BioNTech SE |
NCT02410733 (2023) |
134
|
| Immune-tethered lipoplex nanoparticle (ILN) biochip |
|
|
Diffuse large cell B-lymphoma |
NA |
Recruiting |
Ohio State University Comprehensive Cancer Centre |
NCT03656835 (2022) |
135
|
Delivery challenges and solutions in genosome therapeutics
The clinical translation of genosome-based therapeutics faces significant hurdles related to delivery routes, each presenting unique limitations. Intravenous (i.v.) administration, while widely used, is hampered by the accelerated blood clearance (ABC) phenomenon, where repeated dosing triggers immune-mediated clearance of LNPs, reducing therapeutic efficacy.136 Additionally, passive liver sequestration, driven by endogenous opsonisation, limits extrahepatic targeting, necessitating strategies such as surface charge modulation or pre-dosing with empty LNPs to mitigate undesired biodistribution. Localised delivery, particularly in solid tumours like pancreatic cancer, encounters anatomical barriers such as dense stromal tissue and hypovascularisation, which impede nanoparticle penetration. Emerging solutions, including convection-enhanced delivery and stromal disruption agents, are being explored to enhance tumour accessibility. Inhaled genosomes, though promising for respiratory diseases, face rapid clearance by the mucociliary mechanism and alveolar macrophages, prompting innovations in mucoadhesive coating and sustained-release formulations to prolong lung residency.
Beyond route-specific challenges, technical hurdles in formulation design and manufacturing further complicate genosome development. PEGylation, a common strategy to prolong circulation, paradoxically induces anti-PEG antibodies in up to 40% of patients, leading to hypersensitivity reactions and reduced efficacy upon repeat dosing.137,138 Alternative stealth coatings, such as polyzwitterions and polysarcosine, are under investigation to circumvent immunogenicity while maintaining pharmacokinetic benefits. Scalability remains another critical bottleneck, as batch-to-batch variability in LNP size, encapsulation efficiency, and stability can compromise clinical outcomes. Quality-by-Design (QbD) approaches, coupled with microfluidic manufacturing, are being adopted to enhance reproducibility, yet regulatory alignment on critical quality attributes is still evolving.
Active targeting strategies, though promising, require precise optimisation of ligand density to balance binding avidity and systemic clearance. For instance, folate and RGD peptide-decorated genosomes have demonstrated enhanced tumour accumulation in preclinical models, but excessive ligand loading can trigger off-target uptake or aggregation.139 Recent advances in computational modelling and high-throughput screening are refining ligand-conjugation protocols to achieve optimal targeting efficiency. Furthermore, the integration of stimuli-responsive linkers aims to improve site-specific payload release. Collectively, addressing these delivery challenges and regulatory science will be pivotal for realising the full therapeutic potential of genosomes in diverse clinical settings.
Translational roadmap for genosome therapeutics
The successful clinical translation of genosome-based therapies requires robust monitoring technologies to evaluate biodistribution and therapeutic efficacy. Recent advances in molecular imaging have enabled real-time tracking of genosomes using positron emission tomography (PET) with 64Cu-labelled nanoparticles, providing critical insights into their pharmacokinetics and tissue accumulation patterns.140,141 Additionally, cell-free RNA (cfRNA) in liquid biopsies has emerged as a promising biomarker for assessing delivery efficiency and target engagement, offering a non-invasive approach to monitor treatment response. These innovations in therapeutic monitoring are complemented by developments in bioanalytical methods, including advanced spectroscopy and chromatography techniques, which enhance the characterisation of genosome formulations and their biological interactions. Together, these tools are paving the way for more precise and personalised therapeutic regimens.142
From a regulatory perspective, the approval pathway for genosome therapies is shaped by evolving guidelines from the FDA and EMA, which specify CQAs such as particle size, polydispersity index, and encapsulation efficiency. Notably, regulatory classifications differ significantly between CRISPR-based gene editing and siRNA/mRNA therapies, with the former facing more stringent scrutiny due to permanent genomic modifications. Furthermore, safety assessments require comprehensive chemistry, manufacturing, and controls (CMC) data, particularly for lipid excipients, to address potential immunogenicity and toxicity concerns. Harmonising these regulatory standards across jurisdictions remains a challenge, necessitating ongoing dialogue between developers and agencies to streamline the approval process while ensuring patient safety.143
The patent landscape for genosome technologies reflects rapid innovation in delivery systems and therapeutic applications. Recent filings highlight advancements in ionizable lipid designs, targeted ligands, and scalable manufacturing processes, with a growing emphasis on modular platforms that can be adapted for multiple disease indications. However, intellectual property disputes and overlapping claims pose potential barriers to commercialisation, underscoring the need for clear strategies early in development. Table 7 summarises key recent patents in the field, illustrating the diversity of technological approaches and their assignees. As the field matures, collaboration between academia, industry, and regulators will be essential to translate these innovations into clinically viable therapies that address unmet medical needs.
Table 7.
Recent patent works on lipoplexes
|
Patent No.
|
Filing country
|
Title of work
|
Granted year
|
Ref
|
| US10705085B2 |
United States |
Tethered lipoplex nanoparticle biochips and methods of use |
2020 |
144
|
| RU2671857C1 |
Russia |
New method for production of lipoplex for local introduction and anti-tumor medication that uses such lipoplex |
2018 |
145
|
| RU2784928C2 |
Russia |
Preparation and storage of liposomal RNA compositions suitable for therapy |
2022 |
146
|
| EP3427723B1 |
European Patent Office |
RNA formulation for immunotherapy |
2020 |
147
|
| JP6980230B2 |
Japan |
New branched chain amphipathic lipids |
2021 |
148
|
| JP2018531239A6 |
Japan |
Novel branched-chain amphiphilic lipids |
2021 |
149
|
| EP2998289B1 |
European Patent Office |
Compounds for targeting drug delivery and enhancing siRNA activity |
2019 |
150
|
| JP6383480B2 |
Japan |
Amine-containing transfection reagents and methods for producing and using the same |
2018 |
151
|
| US11124582B2 |
United States |
FLT3L-FC fusion proteins |
2021 |
152
|
| US10662060B2 |
United States |
Manufacture of lipid-based nanoparticles using a dual asymmetric centrifuge |
2020 |
153
|
| JP6905469B2 |
Japan |
Superbranched polymers and polyplexes, and DNA or RNA delivery systems containing them |
2021 |
154
|
| KR102264820B1 |
South Korea |
Stable formulations of lipids and liposomes |
2021 |
155
|
Future perspectives
Genosome technology has emerged as a transformative platform for nucleic acid delivery, offering distinct advantages in biocompatibility, scalability, and structural versatility compared to viral vectors. However, several critical challenges must be addressed to bridge the gap between preclinical promise and clinical translation. A fundamental limitation remains the incomplete characterisation of genosome-biological system interactions, particularly regarding cellular internalisation mechanisms, intracellular trafficking patterns, and endosomal escape efficiency. Advanced molecular imaging modalities, including single-particle tracking microscopy and super-resolution imaging techniques, coupled with high-throughput screening platforms, could provide unprecedented insights into these processes at nanometer resolution. Such fundamental understanding will enable the rational design of next-generation formulations with enhanced delivery efficiency.
The expanding therapeutic potential of RNA-based medicines presents both opportunities and formulation challenges. While genosomes have demonstrated success with single oligonucleotide delivery, the co-encapsulation of multiple RNA species (e.g. siRNA-mRNA combinations) remains technically demanding. Emerging microfluidic production platforms and AI-driven formulation algorithms show particular promise for optimising these complex delivery systems. From a translational perspective, critical pharmaceutical challenges, including long-term stability, lyophilisation compatibility, and prevention of particle aggregation, require systematic investigation. The phenomenon of ABC observed with PEGylated formulations necessitates exploration of alternative stealth technologies, such as zwitterionic polymers or polyglycerol coatings, to enable repeated administration regimens.
Structural optimisation represents a crucial development pathway, with particular emphasis on chemically-defined, biodegradable lipid systems. The design of ionisable lipids with precisely tuned pKa values could significantly improve pH-responsive behaviour and endosomal escape kinetics. Complementary advances in helper lipid chemistry, including the development of zwitterionic phospholipid analogues, may enhance both formulation stability and intracellular delivery efficiency. Targeted delivery strategies continue to evolve, with multifunctional ligand systems (e.g. transferrin-folate conjugates) showing enhanced specificity for challenging therapeutic targets like the BBB. The integration of stimulus-responsive elements (pH, enzyme or redox-sensitive linkers) could further enable disease site-specific activation of therapeutic payloads.
From a manufacturing perspective, the transition to GMP-compliant production using continuous-flow microfluidic systems will be essential for clinical-scale implementation. This must be accompanied by the development of robust analytical characterisation methods to ensure batch-to-batch reproducibility. Comprehensive safety assessment protocols need to address potential immunogenicity concerns and establish detailed biodistribution profiles across relevant disease models. The field would benefit from harmonised regulatory guidelines specific to lipid-based gene delivery systems, covering aspects from physicochemical characterisation to clinical evaluation criteria.
In conclusion, while significant challenges remain, the continued evolution of genosome technology holds tremendous promise for addressing unmet needs across diverse therapeutic areas, including oncology, neurodegenerative disorders, and genetic diseases. By systematically addressing current limitations in formulation science, delivery efficiency, and manufacturing scalability, genosome-based therapies may soon achieve their potential as clinically transformative modalities. The coming decade will likely witness exciting advances as these sophisticated delivery systems transition from laboratory innovation to clinical reality, potentially revolutionising the field of gene therapy.
Ethical and translational considerations
As genosome therapies advance toward clinical application, several ethical considerations merit careful deliberation. These include equitable access to advanced therapies, long-term monitoring of potential off-target effects, and appropriate patient selection criteria. The scientific community must proactively address these concerns through transparent research practices and collaborative engagement with regulatory bodies. Establishing international consensus on manufacturing standards and quality control parameters will be crucial for ensuring both patient safety and therapeutic efficacy. Furthermore, interdisciplinary collaboration spanning lipid chemistry, molecular biology, pharmaceutical sciences, and clinical medicine will be essential to fully realise the potential of genosome technology.
Conclusion
Genosomes have established themselves as a structurally refined category of non-viral delivery systems, demonstrating remarkable adaptability for nucleic acid therapeutics. These lipid-based nanocarriers, incorporating cationic lipids, helper phospholipids, and surface-modified components, provide effective genetic material protection while enabling crucial biological interactions including cellular uptake, endosomal membrane disruption, and intracellular transport. Their superior safety characteristics, notably diminished immunogenic responses and cellular toxicity relative to viral vectors, make them particularly suitable for treating various pathological conditions spanning oncological, neurological, and genetic disorders. While considerable progress has been achieved, several translational hurdles remain to be overcome. Persistent challenges include optimising gene transfer efficiency, enhancing tissue-specific delivery precision, and resolving formulation stability concerns. Future research directions should emphasise intelligent lipid design with environment-responsive properties, development of composite delivery platforms, and refinement of nucleic acid encapsulation methodologies. Incorporation of molecular targeting ligands combined with advanced screening technologies will facilitate the creation of customised therapeutic solutions. For successful clinical translation, three pivotal elements require attention: implementation of scalable production protocols, comprehensive safety evaluation, and establishment of appropriate regulatory guidelines. Emerging data indicate that genosomes possess the potential to become a transformative gene therapy platform, combining favourable safety profiles with the ability to overcome critical delivery obstacles. Continued innovation in this field may provide solutions for numerous unmet medical needs across multiple therapeutic areas, representing a significant advancement in nucleic-acid-based medicine.
Competing Interests
The authors report no financial or any other conflicts of interest in this work.
Ethical Approval
Not applicable.
Acknowledgements
All authors have contributed substantially to the work’s design, performance, analysis, and presentation. The authors would like to express their sincere gratitude to Dr. Sabitha M, principal, Amrita School of Pharmacy, Amrita Vishwa Vidyapeetham, for the constant encouragement, support, and valuable guidance throughout this study.
References
- Vijayan V, Shefrin S, Sreelaxmi C, Nair SC. Aquasomes: an excellent and promising system for novel drug delivery. Int J Pharm Res 2018; 10(2):207-15. [ Google Scholar]
- Collins M, Thrasher A. Gene therapy: progress and predictions. Proc Biol Sci 2015; 282(1821):20143003. doi: 10.1098/rspb.2014.3003 [Crossref] [ Google Scholar]
- Hattori Y. Progress in the development of lipoplex and polyplex modified with anionic polymer for efficient gene delivery. J Genet Med Gene Ther 2017; 1(1):3-18. doi: 10.29328/journal.jgmgt.1001002 [Crossref] [ Google Scholar]
- Raju K, Gayathri PS, Krishnakumar G, Nair SC. Vesosomes: new prospects in multi compartment vesicular drug delivery system. Int J Pharm Res 2020; 12(1):869-77. doi: 10.31838/ijpr/2020.12.01.086 [Crossref] [ Google Scholar]
- Wang H, Jiang Y, Peng H, Chen Y, Zhu P, Huang Y. Recent progress in microRNA delivery for cancer therapy by non-viral synthetic vectors. Adv Drug Deliv Rev 2015; 81:142-60. doi: 10.1016/j.addr.2014.10.031 [Crossref] [ Google Scholar]
- Roberts TC, Ezzat K, El Andaloussi S, Weinberg MS. Synthetic SiRNA delivery: progress and prospects. Methods Mol Biol 2016; 1364:291-310. doi: 10.1007/978-1-4939-3112-5_23 [Crossref] [ Google Scholar]
- Aslam Abdul Rahiman CA, Krishnan K, Sreelekshmi AS, Arjun KK, Nair SC. Novasome: a pioneering advancement in vesicular drug delivery. Int J Appl Pharm 2021; 13(1):59-64. doi: 10.22159/ijap.2021v13i1.39528 [Crossref] [ Google Scholar]
- Young LS, Searle PF, Onion D, Mautner V. Viral gene therapy strategies: from basic science to clinical application. J Pathol 2006; 208(2):299-318. doi: 10.1002/path.1896 [Crossref] [ Google Scholar]
- Morshed M, Chowdhury EH. Gene delivery and clinical applications. In: Narayan R, ed. Encyclopedia of Biomedical Engineering. Vol 1. Elsevier; 2019. p. 345-51. doi: 10.1016/b978-0-12-801238-3.99883-0.
- Ma B, Zhang S, Jiang H, Zhao B, Lv H. Lipoplex morphologies and their influences on transfection efficiency in gene delivery. J Control Release 2007; 123(3):184-94. doi: 10.1016/j.jconrel.2007.08.022 [Crossref] [ Google Scholar]
- Shende P, Ture N, Gaud RS, Trotta F. Lipid- and polymer-based plexes as therapeutic carriers for bioactive molecules. Int J Pharm 2019; 558:250-60. doi: 10.1016/j.ijpharm.2018.12.085 [Crossref] [ Google Scholar]
- Mary Jose A, Lakshmi VU, Gayathri S, Nair SC. Discosomes: a futuristic upheaval in vesicular drug delivery. Int J Appl Pharm 2021; 13(6):41-6. doi: 10.22159/ijap.2021v13i6.42008 [Crossref] [ Google Scholar]
- Pan X, Veroniaina H, Su N, Sha K, Jiang F, Wu Z. Applications and developments of gene therapy drug delivery systems for genetic diseases. Asian J Pharm Sci 2021; 16(6):687-703. doi: 10.1016/j.ajps.2021.05.003 [Crossref] [ Google Scholar]
- Saudagar NR, Boraste SS, Shinkar DM, Pingale PL, Amrutkar SV. Gene-specific drug delivery system: an art of war. BiosciBiotechnol Res Asia 2022; 19(2):459-71. doi: 10.13005/bbra/3000 [Crossref] [ Google Scholar]
- Syyam A, Nawaz A, Ijaz A, Sajjad U, Fazil A, Irfan S. Adenovirus vector system: construction, history and therapeutic applications. Biotechniques 2022; 73(6):297-305. doi: 10.2144/btn-2022-0051 [Crossref] [ Google Scholar]
- Weber T. Anti-AAV antibodies in AAV gene therapy: current challenges and possible solutions. Front Immunol 2021; 12:658399. doi: 10.3389/fimmu.2021.658399 [Crossref] [ Google Scholar]
- Zhao Z, Anselmo AC, Mitragotri S. Viral vector-based gene therapies in the clinic. BioengTransl Med 2022; 7(1):e10258. doi: 10.1002/btm2.10258 [Crossref] [ Google Scholar]
- Hald Albertsen C, Kulkarni JA, Witzigmann D, Lind M, Petersson K, Simonsen JB. The role of lipid components in lipid nanoparticles for vaccines and gene therapy. Adv Drug Deliv Rev 2022; 188:114416. doi: 10.1016/j.addr.2022.114416 [Crossref] [ Google Scholar]
- Salameh JW, Zhou L, Ward SM, Santa Chalarca CF, Emrick T, Figueiredo ML. Polymer-mediated gene therapy: recent advances and merging of delivery techniques. Wiley Interdiscip Rev NanomedNanobiotechnol 2020; 12(2):e1598. doi: 10.1002/wnan.1598 [Crossref] [ Google Scholar]
- Liu Q, Kim YJ, Im GB, Zhu J, Wu Y, Liu Y. Inorganic nanoparticles applied as functional therapeutics. Adv Funct Mater 2021; 31(12):2008171. doi: 10.1002/adfm.202008171 [Crossref] [ Google Scholar]
- Taylor RE, Zahid M. Cell penetrating peptides, novel vectors for gene therapy. Pharmaceutics 2020; 12(3):225. doi: 10.3390/pharmaceutics12030225 [Crossref] [ Google Scholar]
- Su C, Lin D, Huang X, Feng J, Jin A, Wang F. Developing hydrogels for gene therapy and tissue engineering. J Nanobiotechnology 2024; 22(1):182. doi: 10.1186/s12951-024-02462-z [Crossref] [ Google Scholar]
- Karim ME, Shetty J, Islam RA, Kaiser A, Bakhtiar A, Chowdhury EH. Strontium sulfite: a new pH-responsive inorganic nanocarrier to deliver therapeutic siRNAs to cancer cells. Pharmaceutics 2019; 11(2):89. doi: 10.3390/pharmaceutics11020089 [Crossref] [ Google Scholar]
- Kong L, Alves CS, Hou W, Qiu J, Möhwald H, Tomás H. RGD peptide-modified dendrimer-entrapped gold nanoparticles enable highly efficient and specific gene delivery to stem cells. ACS Appl Mater Interfaces 2015; 7(8):4833-43. doi: 10.1021/am508760w [Crossref] [ Google Scholar]
- Davis ME, Zuckerman JE, Choi CH, Seligson D, Tolcher A, Alabi CA. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 2010; 464(7291):1067-70. doi: 10.1038/nature08956 [Crossref] [ Google Scholar]
- Adams D, Gonzalez-Duarte A, O’Riordan WD, Yang CC, Ueda M, Kristen AV. Patisiran, an RNAi therapeutic, for hereditary transthyretin amyloidosis. N Engl J Med 2018; 379(1):11-21. doi: 10.1056/NEJMoa1716153 [Crossref] [ Google Scholar]
- Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell 2009; 136(2):215-33. doi: 10.1016/j.cell.2009.01.002 [Crossref] [ Google Scholar]
- Ambros V. MicroRNA pathways in flies and worms: growth, death, fat, stress, and timing. Cell 2003; 113(6):673-6. doi: 10.1016/s0092-8674(03)00428-8 [Crossref] [ Google Scholar]
- Ha M, Kim VN. Regulation of microRNA biogenesis. Nat Rev Mol Cell Biol 2014; 15(8):509-24. doi: 10.1038/nrm3838 [Crossref] [ Google Scholar]
- Crooke ST. Molecular mechanisms of antisense oligonucleotides. Nucleic Acid Ther 2017; 27(2):70-7. doi: 10.1089/nat.2016.0656 [Crossref] [ Google Scholar]
- Evers MM, Toonen LJ, van Roon-Mom WM. Antisense oligonucleotides in therapy for neurodegenerative disorders. Adv Drug Deliv Rev 2015; 87:90-103. doi: 10.1016/j.addr.2015.03.008 [Crossref] [ Google Scholar]
- Frank-Kamenetsky M, Grefhorst A, Anderson NN, Racie TS, Bramlage B, Akinc A. Therapeutic RNAi targeting PCSK9 acutely lowers plasma cholesterol in rodents and LDL cholesterol in nonhuman primates. Proc Natl Acad Sci U S A 2008; 105(33):11915-20. doi: 10.1073/pnas.0805434105 [Crossref] [ Google Scholar]
- Wijaya YO, Niba ET, Nishio H, Okamoto K, Awano H, Saito T. High concentration or combined treatment of antisense oligonucleotides for spinal muscular atrophy perturbed SMN2 splicing in patient fibroblasts. Genes (Basel) 2022; 13(4):685. doi: 10.3390/genes13040685 [Crossref] [ Google Scholar]
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012; 337(6096):816-21. doi: 10.1126/science.1225829 [Crossref] [ Google Scholar]
- Doudna JA, Charpentier E. Genome editing The new frontier of genome engineering with CRISPR-Cas9. Science 2014; 346(6213):1258096. doi: 10.1126/science.1258096 [Crossref] [ Google Scholar]
- Hsu PD, Lander ES, Zhang F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014; 157(6):1262-78. doi: 10.1016/j.cell.2014.05.010 [Crossref] [ Google Scholar]
- Baumann M. CRISPR/Cas9 genome editing–new and old ethical issues arising from a revolutionary technology. NanoEthics 2016; 10(2):139-59. doi: 10.1007/s11569-016-0259-0 [Crossref] [ Google Scholar]
- Friedrich M, Aigner A. Therapeutic siRNA: state-of-the-art and future perspectives. BioDrugs 2022; 36(5):549-71. doi: 10.1007/s40259-022-00549-3 [Crossref] [ Google Scholar]
- Diener C, Keller A, Meese E. Emerging concepts of miRNA therapeutics: from cells to clinic. Trends Genet 2022; 38(6):613-26. doi: 10.1016/j.tig.2022.02.006 [Crossref] [ Google Scholar]
- Chakkyarath V. Applications of advanced ASOs, RNA mimics, and others in RNA-based therapeutics. In: Sharma NK, Chand Dakal T, Maurya PK, Wadhwa G, eds. RNA-Based Cancer Therapeutics: Techniques, Applications and Challenges in Prognosis. Singapore: Springer; 2025. p. 35-62. doi: 10.1007/978-981-96-3456-9_3.
- Huang J, Zhou Y, Li J, Lu A, Liang C. CRISPR/Cas systems: delivery and application in gene therapy. Front BioengBiotechnol 2022; 10:942325. doi: 10.3389/fbioe.2022.942325 [Crossref] [ Google Scholar]
- Jayakumar R, Prabaharan M, Muzzarelli RA. Chitosan for Biomaterials I. Vol 243. Heidelberg: Springer; 2011.
- Ostróżka-Cieślik A, Sarecka-Hujar B. The use of nanotechnology in modern pharmacotherapy. In: Grumezescu AM, ed. Multifunctional Systems for Combined Delivery, Biosensing and Diagnostics. Elsevier; 2017. p. 139-58. doi: 10.1016/b978-0-323-52725-5.00007-1.
- Dan N. Structure and kinetics of synthetic, lipid-based nucleic acid carriers: lipoplexes. In: Grumezescu AM, ed. Lipid Nanocarriers for Drug Targeting. William Andrew Publishing; 2018. p. 529-62. doi: 10.1016/b978-0-12-813687-4.00014-1.
- Jayakumar R, Menon D, Manzoor K, Nair SV, Tamura H. Biomedical applications of chitin and chitosan-based nanomaterials—a short review. CarbohydrPolym 2010; 82(2):227-32. doi: 10.1016/j.carbpol.2010.04.074 [Crossref] [ Google Scholar]
- Ewert K, Evans HM, Ahmad A, Slack NL, Lin AJ, Martin-Herranz A. Lipoplex structures and their distinct cellular pathways. Adv Genet 2005; 53:119-55. doi: 10.1016/s0065-2660(05)53005-0 [Crossref] [ Google Scholar]
- Kopatz I, Remy JS, Behr JP. A model for non-viral gene delivery: through syndecan adhesion molecules and powered by actin. J Gene Med 2004; 6(7):769-76. doi: 10.1002/jgm.558 [Crossref] [ Google Scholar]
- Mislick KA, Baldeschwieler JD. Evidence for the role of proteoglycans in cation-mediated gene transfer. Proc Natl Acad Sci U S A 1996; 93(22):12349-54. doi: 10.1073/pnas.93.22.12349 [Crossref] [ Google Scholar]
- Elouahabi A, Ruysschaert JM. Formation and intracellular trafficking of lipoplexes and polyplexes. Mol Ther 2005; 11(3):336-47. doi: 10.1016/j.ymthe.2004.12.006 [Crossref] [ Google Scholar]
- Zuhorn IS, Hoekstra D. On the mechanism of cationic amphiphile-mediated transfection. To fuse or not to fuse: is that the question?. J Membr Biol 2002; 189(3):167-79. doi: 10.1007/s00232-002-1015-7 [Crossref] [ Google Scholar]
- Lasic DD, Strey H, Stuart MC, Podgornik R, Frederik PM. The structure of DNA−liposome complexes. J Am Chem Soc 1997; 119(4):832-3. doi: 10.1021/ja962713g [Crossref] [ Google Scholar]
- Rädler JO, Koltover I, Salditt T, Safinya CR. Structure of DNA-cationic liposome complexes: DNA intercalation in multilamellar membranes in distinct interhelical packing regimes. Science 1997; 275(5301):810-4. doi: 10.1126/science.275.5301.810 [Crossref] [ Google Scholar]
- Koltover I, Salditt T, Rädler JO, Safinya CR. An inverted hexagonal phase of cationic liposome-DNA complexes related to DNA release and delivery. Science 1998; 281(5373):78-81. doi: 10.1126/science.281.5373.78 [Crossref] [ Google Scholar]
- Ewert KK, Evans HM, Zidovska A, Bouxsein NF, Ahmad A, Safinya CR. A columnar phase of dendritic lipid-based cationic liposome-DNA complexes for gene delivery: hexagonally ordered cylindrical micelles embedded in a DNA honeycomb lattice. J Am Chem Soc 2006; 128(12):3998-4006. doi: 10.1021/ja055907h [Crossref] [ Google Scholar]
- Dan N, Danino D. Structure and kinetics of lipid-nucleic acid complexes. Adv Colloid Interface Sci 2014; 205:230-9. doi: 10.1016/j.cis.2014.01.013 [Crossref] [ Google Scholar]
- Barreleiro PC, Lindman B. The kinetics of DNA−cationic vesicle complex formation. J Phys Chem B 2003; 107(25):6208-13. doi: 10.1021/jp0277107 [Crossref] [ Google Scholar]
- Barreleiro PC, May RP, Lindman B. Mechanism of formation of DNA-cationic vesicle complexes. Faraday Discuss 2003; 122:191-201. doi: 10.1039/b200796g [Crossref] [ Google Scholar]
- Kennedy MT, Pozharski EV, Rakhmanova VA, MacDonald RC. Factors governing the assembly of cationic phospholipid-DNA complexes. Biophys J 2000; 78(3):1620-33. doi: 10.1016/s0006-3495(00)76714-2 [Crossref] [ Google Scholar]
- Pozharski EV, MacDonald RC. Single lipoplex study of cationic lipoid-DNA, self-assembled complexes. Mol Pharm 2007; 4(6):962-74. doi: 10.1021/mp700080m [Crossref] [ Google Scholar]
- Simberg D, Danino D, Talmon Y, Minsky A, Ferrari ME, Wheeler CJ. Phase behavior, DNA ordering, and size instability of cationic lipoplexes Relevance to optimal transfection activity. J Biol Chem 2001; 276(50):47453-9. doi: 10.1074/jbc.M105588200 [Crossref] [ Google Scholar]
- Kapoor M, Burgess DJ, Patil SD. Physicochemical characterization techniques for lipid-based delivery systems for siRNA. Int J Pharm 2012; 427(1):35-57. doi: 10.1016/j.ijpharm.2011.09.032 [Crossref] [ Google Scholar]
- Eastman SJ, Siegel C, Tousignant J, Smith AE, Cheng SH, Scheule RK. Biophysical characterization of cationic lipid: DNA complexes. BiochimBiophys Acta 1997; 1325(1):41-62. doi: 10.1016/s0005-2736(96)00242-8 [Crossref] [ Google Scholar]
- Felgner PL, Ringold GM. Cationic liposome-mediated transfection. Nature 1989; 337(6205):387-8. doi: 10.1038/337387a0 [Crossref] [ Google Scholar]
- Zuhorn IS, Oberle V, Visser WH, Engberts JB, Bakowsky U, Polushkin E. Phase behavior of cationic amphiphiles and their mixtures with helper lipid influences lipoplex shape, DNA translocation, and transfection efficiency. Biophys J 2002; 83(4):2096-108. doi: 10.1016/s0006-3495(02)73970-2 [Crossref] [ Google Scholar]
- Ruysschaert JM, Elouahabi A, Willeaume V, Huez G, Fuks R, Vandenbranden M. A novel cationic amphiphile for transfection of mammalian cells. BiochemBiophys Res Commun 1994; 203(3):1622-8. doi: 10.1006/bbrc.1994.2372 [Crossref] [ Google Scholar]
- Sakurai F, Inoue R, Nishino Y, Okuda A, Matsumoto O, Taga T. Effect of DNA/liposome mixing ratio on the physicochemical characteristics, cellular uptake and intracellular trafficking of plasmid DNA/cationic liposome complexes and subsequent gene expression. J Control Release 2000; 66(2-3):255-69. doi: 10.1016/s0168-3659(99)00280-1 [Crossref] [ Google Scholar]
- Luiz MT, Dutra JA, Tofani LB, de Araújo JT, Di Filippo LD, Marchetti JM. Targeted liposomes: a nonviral gene delivery system for cancer therapy. Pharmaceutics 2022; 14(4):821. doi: 10.3390/pharmaceutics14040821 [Crossref] [ Google Scholar]
- Fujii S, Nishimura T, Sakurai K. Thermodynamics of lipoplex formation: relationship between the lipid alkyl tail length and thermodynamic functions. Chem Lett 2012; 41(5):501-3. doi: 10.1246/cl.2012.501 [Crossref] [ Google Scholar]
- Janich C, Wölk C, Erdmann F, Groth T, Brezesinski G, Dobner B. Composites of malonic acid diamides and phospholipids--impact of lipoplex stability on transfection efficiency. J Control Release 2015; 220(Pt A):295-307. doi: 10.1016/j.jconrel.2015.10.045 [Crossref] [ Google Scholar]
- Józkowicz A, Dulak J. New strategies for application of plasmid and viral vectors in gene therapy. Biotechnologia 2007; 78(3):7-21. [ Google Scholar]
- Rakhmanova VA, Mcintosh TJ, Macdonald RC. Effects of dioleoylphosphatidylethanolamine on the activity and structure of O-alkyl phosphatidylcholine-DNA transfection complexes. Cell Mol Biol Lett 2000; 5(1):51-65. [ Google Scholar]
- Wang L, Yoshida J, Ogata N, Sasaki S, Kajiyama T. Self-assembled supramolecular films derived from marine deoxyribonucleic acid (DNA)−cationic surfactant complexes: large-scale preparation and optical and thermal properties. Chem Mater 2001; 13(4):1273-81. doi: 10.1021/cm000869g [Crossref] [ Google Scholar]
- Buck J, Mueller D, Mettal U, Ackermann M, Grisch-Chan HM, Thöny B. Improvement of DNA vector delivery of DOTAP lipoplexes by short-chain aminolipids. ACS Omega 2020; 5(38):24724-32. doi: 10.1021/acsomega.0c03303 [Crossref] [ Google Scholar]
- Yavuz A, Coiffier C, Garapon C, Gurcan S, Monge C, Exposito JY. DLin-MC3-containing mRNA lipid nanoparticles induce an antibody Th2-biased immune response polarization in a delivery route-dependent manner in mice. Pharmaceutics 2023; 15(3):1009. doi: 10.3390/pharmaceutics15031009 [Crossref] [ Google Scholar]
- Banerji A, Wickner PG, Saff R, Stone CA Jr, Robinson LB, Long AA. mRNA vaccines to prevent COVID-19 disease and reported allergic reactions: current evidence and suggested approach. J Allergy Clin Immunol Pract 2021; 9(4):1423-37. doi: 10.1016/j.jaip.2020.12.047 [Crossref] [ Google Scholar]
- Hattori Y, Tamaki K, Ozaki KI, Kawano K, Onishi H. Optimized combination of cationic lipids and neutral helper lipids in cationic liposomes for siRNA delivery into the lung by intravenous injection of siRNA lipoplexes. J Drug Deliv Sci Technol 2019; 52:1042-50. doi: 10.1016/j.jddst.2019.06.016 [Crossref] [ Google Scholar]
- Shmendel EV, Puchkov PA, Maslov MA. Design of folate-containing liposomal nucleic acid delivery systems for antitumor therapy. Pharmaceutics 2023; 15(5):1400. doi: 10.3390/pharmaceutics15051400 [Crossref] [ Google Scholar]
- Zabner J, Fasbender AJ, Moninger T, Poellinger KA, Welsh MJ. Cellular and molecular barriers to gene transfer by a cationic lipid. J Biol Chem 1995; 270(32):18997-9007. doi: 10.1074/jbc.270.32.18997 [Crossref] [ Google Scholar]
- Zhdanov RI, Podobed OV, Vlassov VV. Cationic lipid-DNA complexes-lipoplexes-for gene transfer and therapy. Bioelectrochemistry 2002; 58(1):53-64. doi: 10.1016/s1567-5394(02)00132-9 [Crossref] [ Google Scholar]
- Wrobel I, Collins D. Fusion of cationic liposomes with mammalian cells occurs after endocytosis. BiochimBiophys Acta 1995; 1235(2):296-304. doi: 10.1016/0005-2736(95)80017-a [Crossref] [ Google Scholar]
- Degors IMS, Wang C, Rehman ZU, Zuhorn IS. Carriers break barriers in drug delivery: endocytosis and endosomal escape of gene delivery vectors. Acc Chem Res 2019; 52(7):1750-60. doi: 10.1021/acs.accounts.9b00177 [Crossref] [ Google Scholar]
- Liberali P, Rämö P, Pelkmans L. Protein kinases: starting a molecular-systems view of endocytosis. Annu Rev Cell Dev Biol 2008; 24:501-23. doi: 10.1146/annurev.cellbio.041008.145637 [Crossref] [ Google Scholar]
- Rozema DB. The chemistry of oligonucleotide delivery. Annu Rep Med Chem 2017; 50:17-59. doi: 10.1016/bs.armc.2017.07.003 [Crossref] [ Google Scholar]
- Pichon C, Billiet L, Midoux P. Chemical vectors for gene delivery: uptake and intracellular trafficking. CurrOpinBiotechnol 2010; 21(5):640-5. doi: 10.1016/j.copbio.2010.07.003 [Crossref] [ Google Scholar]
- Simões S, Slepushkin V, Pires P, Gaspar R, Pedroso de Lima MC, Düzgüneş N. Human serum albumin enhances DNA transfection by lipoplexes and confers resistance to inhibition by serum. BiochimBiophys Acta 2000; 1463(2):459-69. doi: 10.1016/s0005-2736(99)00238-2 [Crossref] [ Google Scholar]
- Fang Y, Yang J. Two-dimensional condensation of DNA molecules on cationic lipid membranes. J Phys Chem B 1997; 101(3):441-9. doi: 10.1021/jp962382u [Crossref] [ Google Scholar]
- Haghighi E, Abolmaali SS, Dehshahri A, Mousavi Shaegh SA, Azarpira N, Tamaddon AM. Navigating the intricate in-vivo journey of lipid nanoparticles tailored for the targeted delivery of RNA therapeutics: a quality-by-design approach. J Nanobiotechnology 2024; 22(1):710. doi: 10.1186/s12951-024-02972-w [Crossref] [ Google Scholar]
- Čechová P, Paloncýová M, Šrejber M, Otyepka M. Mechanistic insights into interactions between ionizable lipid nanodroplets and biomembranes. J Biomol Struct Dyn 2024:1-11. doi: 10.1080/07391102.2024.2329307.
- Aliakbarinodehi N, Gallud A, Mapar M, Wesén E, Heydari S, Jing Y. Interaction kinetics of individual mRNA-containing lipid nanoparticles with an endosomal membrane mimic: dependence on pH, protein corona formation, and lipoprotein depletion. ACS Nano 2022; 16(12):20163-73. doi: 10.1021/acsnano.2c04829 [Crossref] [ Google Scholar]
- Ma Y, Li S, Lin X, Chen Y. A perspective of lipid nanoparticles for RNA delivery. Exploration (Beijing) 2024; 4(6):20230147. doi: 10.1002/exp.20230147 [Crossref] [ Google Scholar]
- Gandek TB, van der Koog L, Nagelkerke A. A comparison of cellular uptake mechanisms, delivery efficacy, and intracellular fate between liposomes and extracellular vesicles. Adv Healthc Mater 2023; 12(25):e2300319. doi: 10.1002/adhm.202300319 [Crossref] [ Google Scholar]
- Kulkarni JA, Witzigmann D, Leung J, Tam YY, Cullis PR. On the role of helper lipids in lipid nanoparticle formulations of siRNA. Nanoscale 2019; 11(45):21733-9. doi: 10.1039/c9nr09347h [Crossref] [ Google Scholar]
- Mohammadi A, Kudsiova L, Mohd Mustapa MF, Campbell F, Vlaho D, Welser K. The discovery and enhanced properties of trichain lipids in lipopolyplex gene delivery systems. Org Biomol Chem 2019; 17(4):945-57. doi: 10.1039/c8ob02374c [Crossref] [ Google Scholar]
- Seraj S, Lee J, Ahn HJ. Systemic delivery of Eg5 shRNA-expressing plasmids using PEGylated DC-Chol/DOPE cationic liposome: long-term silencing and anticancer effects in vivo. BiochemPharmacol 2019; 166:192-202. doi: 10.1016/j.bcp.2019.05.021 [Crossref] [ Google Scholar]
- Forssen EA, Coulter DM, Proffitt RT. Selective in vivo localization of daunorubicin small unilamellar vesicles in solid tumors. Cancer Res 1992; 52(12):3255-61. [ Google Scholar]
- Tomlinson E, Rolland AP. Controllable gene therapy pharmaceutics of non-viral gene delivery systems. J Control Release 1996; 39(2-3):357-72. doi: 10.1016/0168-3659(95)00166-2 [Crossref] [ Google Scholar]
- Coune PG, Schneider BL, Aebischer P. Parkinson’s disease: gene therapies. Cold Spring HarbPerspect Med 2012; 2(4):a009431. doi: 10.1101/cshperspect.a009431 [Crossref] [ Google Scholar]
- Pandian SR, Vijayakumar KK, Murugesan S, Kunjiappan S. Liposomes: an emerging carrier for targeting Alzheimer’s and Parkinson’s diseases. Heliyon 2022; 8(6):e09575. doi: 10.1016/j.heliyon.2022.e09575 [Crossref] [ Google Scholar]
- Hattori Y, Tamaki K, Sakasai S, Ozaki KI, Onishi H. Effects of PEG anchors in PEGylated siRNA lipoplexes on in vitro gene-silencing effects and siRNA biodistribution in mice. Mol Med Rep 2020; 22(5):4183-96. doi: 10.3892/mmr.2020.11525 [Crossref] [ Google Scholar]
- Nisini R, Poerio N, Mariotti S, De Santis F, Fraziano M. The multirole of liposomes in therapy and prevention of infectious diseases. Front Immunol 2018; 9:155. doi: 10.3389/fimmu.2018.00155 [Crossref] [ Google Scholar]
- Domashenko A, Gupta S, Cotsarelis G. Efficient delivery of transgenes to human hair follicle progenitor cells using topical lipoplex. Nat Biotechnol 2000; 18(4):420-3. doi: 10.1038/74480 [Crossref] [ Google Scholar]
- Abu Lila AS, Uehara Y, Ishida T, Kiwada H. Application of polyglycerol coating to plasmid DNA lipoplex for the evasion of the accelerated blood clearance phenomenon in nucleic acid delivery. J Pharm Sci 2014; 103(2):557-66. doi: 10.1002/jps.23823 [Crossref] [ Google Scholar]
- Grabbe S, Haas H, Diken M, Kranz LM, Langguth P, Sahin U. Translating nanoparticulate-personalized cancer vaccines into clinical applications: case study with RNA-lipoplexes for the treatment of melanoma. Nanomedicine (Lond) 2016; 11(20):2723-34. doi: 10.2217/nnm-2016-0275 [Crossref] [ Google Scholar]
- Santel A, Aleku M, Keil O, Endruschat J, Esche V, Fisch G. A novel siRNA-lipoplex technology for RNA interference in the mouse vascular endothelium. Gene Ther 2006; 13(16):1222-34. doi: 10.1038/sj.gt.3302777 [Crossref] [ Google Scholar]
- Rossi SM, Murray T, McDonough L, Kelly H. Loco-regional drug delivery in oncology: current clinical applications and future translational opportunities. Expert Opin Drug Deliv 2021; 18(5):607-23. doi: 10.1080/17425247.2021.1856074 [Crossref] [ Google Scholar]
- Zhang WW, Li L, Li D, Liu J, Li X, Li W. The first approved gene therapy product for cancer Ad-p53 (Gendicine): 12 years in the clinic. Hum Gene Ther 2018; 29(2):160-79. doi: 10.1089/hum.2017.218 [Crossref] [ Google Scholar]
- Karmali PP, Chaudhuri A. Cationic liposomes as non-viral carriers of gene medicines: resolved issues, open questions, and future promises. Med Res Rev 2007; 27(5):696-722. doi: 10.1002/med.20090 [Crossref] [ Google Scholar]
- Vaidya S, Mohod A, Eedara AC, Andugulapati SB, Pabbaraja S. Synthesis and characterization of a new cationic lipid: efficient siRNA delivery and anticancer activity of survivin‐siRNA lipoplexes for the treatment of lung and breast cancers. ChemMedChem 2023; 18(16):e202300097. doi: 10.1002/cmdc.202300097 [Crossref] [ Google Scholar]
- Akhter A. Development of a Novel Active Targeting System for siRNA Delivery to Liver Sinusoidal Endothelial Cell in Vivo. Hokkaido University; 2013. doi: 10.14943/doctoral.k11117.
- Andey T, Bora-Singhal N, Chellappan SP, Singh M. Cationic lipoplexes for treatment of cancer stem cell-derived murine lung tumors. Nanomedicine 2019; 18:31-43. doi: 10.1016/j.nano.2019.02.007 [Crossref] [ Google Scholar]
- Kowzun MJ, Rifkin WJ, Borab ZM, Ellison T, Soares MA, Wilson SC. Topical inhibition of PUMA signaling mitigates radiation injury. Wound Repair Regen 2018; 26(6):413-25. doi: 10.1111/wrr.12668 [Crossref] [ Google Scholar]
- Brahmamdam P, Watanabe E, Unsinger J, Chang KC, Schierding W, Hoekzema AS. Targeted delivery of siRNA to cell death proteins in sepsis. Shock 2009; 32(2):131-9. doi: 10.1097/SHK.0b013e318194bcee [Crossref] [ Google Scholar]
- Sugiyama-Nakagiri Y, Akiyama M, Shimizu H. Hair follicle stem cell-targeted gene transfer and reconstitution system. Gene Ther 2006; 13(8):732-7. doi: 10.1038/sj.gt.3302709 [Crossref] [ Google Scholar]
- Zheng W, Xu CH. Innovative approaches and advances for hair follicle regeneration. ACS Biomater Sci Eng 2023; 9(5):2251-76. doi: 10.1021/acsbiomaterials.3c00028 [Crossref] [ Google Scholar]
- Villate-Beitia I, Puras G, Soto-Sánchez C, Agirre M, Ojeda E, Zarate J. Non-viral vectors based on magnetoplexes, lipoplexes and polyplexes for VEGF gene delivery into central nervous system cells. Int J Pharm 2017; 521(1-2):130-40. doi: 10.1016/j.ijpharm.2017.02.016 [Crossref] [ Google Scholar]
- Vera-López KJ, Aranzamendi-Zenteno M, Davila-Del-Carpio G, Nieto-Montesinos R. Using immunoliposomes as carriers to enhance the therapeutic effectiveness of macamide N-3-methoxybenzyl-linoleamide. Neurol Int 2025; 17(3):38. doi: 10.3390/neurolint17030038 [Crossref] [ Google Scholar]
- Pardridge WM. Treatment of Parkinson’s disease with biologics that penetrate the blood-brain barrier via receptor-mediated transport. Front Aging Neurosci 2023; 15:1276376. doi: 10.3389/fnagi.2023.1276376 [Crossref] [ Google Scholar]
- Chen W, Li H, Liu Z, Yuan W. Lipopolyplex for therapeutic gene delivery and its application for the treatment of Parkinson’s disease. Front Aging Neurosci 2016; 8:68. doi: 10.3389/fnagi.2016.00068 [Crossref] [ Google Scholar]
- Jones CH, Chen CK, Ravikrishnan A, Rane S, Pfeifer BA. Overcoming nonviral gene delivery barriers: perspective and future. Mol Pharm 2013; 10(11):4082-98. doi: 10.1021/mp400467x [Crossref] [ Google Scholar]
- Pu Y, Dai XL, Wang Y, Chen Y, Wu C, Zhou X. Hybrid lipoplex boosts neuron-microglia crosstalk for treatment of Alzheimer’s disease through Aβ-targeted-autophagy and ApoE2 gene supplementation. Adv Mater 2025; 37(27):e2418560. doi: 10.1002/adma.202418560 [Crossref] [ Google Scholar]
- Erel-Akbaba G, Akbaba H, Karaman O, Tian T, Tannous BA, Turunc E. Rabies virus-mimicking liposomes for targeted gene therapy in Alzheimer’s disease. Int J Pharm 2025; 668:124962. doi: 10.1016/j.ijpharm.2024.124962 [Crossref] [ Google Scholar]
- Carvalheiro M, Esteves MA, Santos-Mateus D, Lopes RM, Rodrigues MA, Eleutério CV. Hemisynthetic trifluralin analogues incorporated in liposomes for the treatment of leishmanial infections. Eur J Pharm Biopharm 2015; 93:346-52. doi: 10.1016/j.ejpb.2015.04.018 [Crossref] [ Google Scholar]
- Mohmmed A, Dasaradhi PV, Bhatnagar RK, Chauhan VS, Malhotra P. In vivo gene silencing in Plasmodium berghei--a mouse malaria model. BiochemBiophys Res Commun 2003; 309(3):506-11. doi: 10.1016/j.bbrc.2003.08.027 [Crossref] [ Google Scholar]
- Janse CJ, Ramesar J, Waters AP. High-efficiency transfection and drug selection of genetically transformed blood stages of the rodent malaria parasite Plasmodium berghei. Nat Protoc 2006; 1(1):346-56. doi: 10.1038/nprot.2006.53 [Crossref] [ Google Scholar]
- Markov OO, Mironova NL, Maslov MA, Petukhov IA, Morozova NG, Vlassov VV. Novel cationic liposomes provide highly efficient delivery of DNA and RNA into dendritic cell progenitors and their immature offsets. J Control Release 2012; 160(2):200-10. doi: 10.1016/j.jconrel.2011.11.034 [Crossref] [ Google Scholar]
- Tenchov R, Bird R, Curtze AE, Zhou Q. Lipid nanoparticles─from liposomes to mRNA vaccine delivery, a landscape of research diversity and advancement. ACS Nano 2021; 15(11):16982-7015. doi: 10.1021/acsnano.1c04996 [Crossref] [ Google Scholar]
- Van Hoecke L, Roose K, Ballegeer M, Zhong Z, Sanders NN, De Koker S. The opposing effect of type I IFN on the T cell response by non-modified mRNA-lipoplex vaccines is determined by the route of administration. Mol Ther Nucleic Acids 2020; 22:373-81. doi: 10.1016/j.omtn.2020.09.004 [Crossref] [ Google Scholar]
- Dixon S, Kang X, Quan D. Practical guidance for the use of patisiran in the management of polyneuropathy in hereditary transthyretin-mediated amyloidosis. Ther Clin Risk Manag 2023; 19:973-81. doi: 10.2147/tcrm.S361706 [Crossref] [ Google Scholar]
- Thi TT, Suys EJ, Lee JS, Nguyen DH, Park KD, Truong NP. Lipid-based nanoparticles in the clinic and clinical trials: from cancer nanomedicine to COVID-19 vaccines. Vaccines (Basel) 2021; 9(4):359. doi: 10.3390/vaccines9040359 [Crossref] [ Google Scholar]
- Zhang X, Goel V, Attarwala H, Sweetser MT, Clausen VA, Robbie GJ. Patisiran pharmacokinetics, pharmacodynamics, and exposure-response analyses in the phase 3 APOLLO trial in patients with hereditary transthyretin-mediated (hATTR) amyloidosis. J Clin Pharmacol 2020; 60(1):37-49. doi: 10.1002/jcph.1480 [Crossref] [ Google Scholar]
- Lee Y, Jeong M, Park J, Jung H, Lee H. Immunogenicity of lipid nanoparticles and its impact on the efficacy of mRNA vaccines and therapeutics. Exp Mol Med 2023; 55(10):2085-96. doi: 10.1038/s12276-023-01086-x [Crossref] [ Google Scholar]
- Rao DD, Jay C, Wang Z, Luo X, Kumar P, Eysenbach H. Preclinical justification of pbi-shRNA EWS/FLI1 lipoplex (LPX) treatment for Ewing’s sarcoma. Mol Ther 2016; 24(8):1412-22. doi: 10.1038/mt.2016.93 [Crossref] [ Google Scholar]
- Rana S, Maples PB, Senzer N, Nemunaitis J. Stathmin 1: a novel therapeutic target for anticancer activity. Expert Rev Anticancer Ther 2008; 8(9):1461-70. doi: 10.1586/14737140.8.9.1461 [Crossref] [ Google Scholar]
- Sahin U, Oehm P, Derhovanessian E, Jabulowsky RA, Vormehr M, Gold M. An RNA vaccine drives immunity in checkpoint-inhibitor-treated melanoma. Nature 2020; 585(7823):107-12. doi: 10.1038/s41586-020-2537-9 [Crossref] [ Google Scholar]
- McCarthy ÁC. Study of the Roles of Autophagy in Human Follicular and Diffuse Large B-Cell Lymphoma [dissertation]. Queen Mary University of London; 2014.
- Lai C, Li C, Liu M, Qiu Q, Luo X, Liu X. Effect of Kupffer cells depletion on ABC phenomenon induced by Kupffer cells-targeted liposomes. Asian J Pharm Sci 2019; 14(4):455-64. doi: 10.1016/j.ajps.2018.07.004 [Crossref] [ Google Scholar]
- Gaballa SA, Naguib Y, Mady FM, Khaled KA. Polyethylene glycol: properties, applications, and challenges. J Adv Biomed Pharm Sci 2024; 7(1):26-36. doi: 10.21608/jabps.2023.241685.1205 [Crossref] [ Google Scholar]
- d’Avanzo N, Celia C, Barone A, Carafa M, Di Marzio L, Santos HA. Immunogenicity of polyethylene glycol-based nanomedicines: mechanisms, clinical implications and systematic approach. Adv Ther (Weinh) 2020; 3(3):1900170. doi: 10.1002/adtp.201900170 [Crossref] [ Google Scholar]
- Gladkikh DV, Sen Kova AV, Chernikov IV, Kabilova TO, Popova NA, Nikolin VP. Folate-equipped cationic liposomes deliver anti-MDR1-siRNA to the tumor and increase the efficiency of chemotherapy. Pharmaceutics 2021; 13(8):1252. doi: 10.3390/pharmaceutics13081252 [Crossref] [ Google Scholar]
- Forte E, Fiorenza D, Torino E, Costagliola di Polidoro A, Cavaliere C, Netti PA. Radiolabeled PET/MRI nanoparticles for tumor imaging. J Clin Med 2019; 9(1):89. doi: 10.3390/jcm9010089 [Crossref] [ Google Scholar]
- Julius A, Renuka RR, Malakondaiah S, Ramalingam S, Dharmalingam Jothinathan MK, Srinivasan GP. Radiolabeled nanoparticles in multimodal nuclear imaging, diagnostics and therapy. J RadioanalNucl Chem 2025; 334(7):4403-18. doi: 10.1007/s10967-025-10233-9 [Crossref] [ Google Scholar]
- Đorđević S, Gonzalez MM, Conejos-Sánchez I, Carreira B, Pozzi S, Acúrcio RC. Current hurdles to the translation of nanomedicines from bench to the clinic. Drug DelivTransl Res 2022; 12(3):500-25. doi: 10.1007/s13346-021-01024-2 [Crossref] [ Google Scholar]
- Grewal S, Sharma T, Swami R. New era of nanoparticles facilitated co-delivery in cancer therapy: two heads are better than one. J Nanopart Res 2023; 25(9):190. doi: 10.1007/s11051-023-05837-9 [Crossref] [ Google Scholar]
- Lee LJ, Kwak KJ, Yu B, Wu Y, Lee A. US10705085B2 - Tethered Lipoplex Nanoparticle Biochips and Methods of Use. United States: Google Patents; 2020.
- Yoshima K, Ishida A, Fukushima A. RU2671857C1 - New Method for Production of Lipoplex for Local Introduction and Anti-Tumor Medication That Uses Such Lipoplex. Russia: Google Patents; 2018.
- Haas H, Hörner S, Esparza Borquez IH, Hiller TM, Bates F. RU2784928C2 - Preparation and Storage of Liposomal RNA Formulations Suitable for Therapy. Russia: Google Patents; 2018.
- Sahin U, Haas H, Kreiter S, Diken M, Fritz D, Meng M, et al. EP3427723B1 - RNA Formulation for Immunotherapy. European Patent Office; 2020.
- Jaffray A, Cawson-Coulves PA, Afonso H, Montierre D, Gall T, Le T. JP6980230B2 - New Branched Chain Amphipathic Lipids. Japan: Google Patents; 2021.
- Jaffray A, Cawson-Coulves PA, Afonso H, Montierre D, Gall T, Le T. JP2018531239A6 - Novel Branched-Chain Amphiphilic Lipids. Japan: Google Patents; 2021.
- Nagarajan Sridhar C, Karmali P, Payne JE, Hou Z, Gaudette JA, Knopov V, et al. EP2998289B1 - Compounds for Targeting Drug Delivery and Enhancing siRNA Activity. European Patent Office; 2019.
- Yang Z, Angrish P, de Mollerat du Jeu X, Wiederholt K. JP6383480B2 - Amine-Containing Transfection Reagents and Methods for Producing and Using the Same. Japan: Google Patents; 2018.
- Ambrogelly A, Baca M, Carr BA, Chu HM, Hung MS, Kanwar M, et al. US11124582B2 - FLT3L-FC Fusion Proteins. United States: Google Patents; 2021.
- Massing U. US10662060B2 - Manufacture of Lipid-Based Nanoparticles Using a Dual Asymmetric Centrifuge. United States: Google Patents; 2020.
- Ma PX, Zhang X. JP6905469B2 - Superbranched Polymers and Polyplexes, and DNA or RNA Delivery Systems Containing Them. Japan: Google Patents; 2021.
- Haas H, Esparza Borquez IH. KR102264820B1 - Stable Formulations of Lipids and Liposomes. South Korea: Google Patents; 2021.