Advanced pharmaceutical bulletin. 15(3):533-551.
doi: 10.34172/apb.025.45330
Review Article
Therapeutic Potential of Circular RNAs as Targets for Cancer Treatment
Milad Asadi Writing – original draft, Writing – review & editing, 1 
Sanam Sadeghi-Mohammadi Methodology, Writing – original draft, 2
Najibeh Shekari Writing – original draft, 3
Venus Zafari Writing – original draft, 1
Zahra Soleimani Methodology, Writing – original draft, 4
Ufuk Mert Supervision, Writing – original draft, 5
Maryam Seyyedi Supervision, Writing – original draft, 6
Ayse Caner Project administration, Supervision, Writing – original draft, Writing – review & editing, 1, 7, *
Habib Zarredar Project administration, Writing – original draft, Writing – review & editing, 6, *
Author information:
1Department of Basic Oncology, Institute of Health Sciences, Ege University, Izmir, Turkey
2ATMP Department, Breast Cancer Research Center, Motamed Cancer Institute, ACECR, Tehran, Iran
3Department of Immunology, Faculty of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran
4Rahat Breath and Sleep Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
5Atatürk Health Care Vocational School, Ege University, Izmir, Turkey
6Tuberculosis and Lung Diseases Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
7Department of Parasitology, Faculty of Medicine, Ege University, Izmir, Turkey
Abstract
Circular RNAs (circRNAs) are a novel class of non-coding RNAs primarily generated through a back-splicing processes. These molecules exhibit extensive expression across various tissues, indicating their significant role in numerous biological processes, particularly in complex diseases such as cancer. Based on their origin, structure, and biogenesis, circular RNAs are categorized into exonic circRNAs (ecirc-RNAs), circular intronic RNAs (ci-RNAs), or exonic-intronic circRNAs (EIci-RNAs). Due to their covalently closed-loop configuration, it is necessary to develop specialized techniques to study them. CircRNAs are known to function as protein and microRNA sponges, regulate transcription, interact with RNA-binding proteins (RBPs), and, in rare cases, serve as templates for translation. In this review, we provide an overview of circRNA features, biogenesis, and functions. In addition, we summarize molecular methods for studying them and explain their significant roles in malignancies.
Keywords: Circular RNA, Biogenesis, Diagnosis, Prognosis, Cancer
Copyright and License Information
© 2025 The Author (s).
This is an Open Access article distributed under the terms of the Creative Commons Attribution (CC BY), which permits unrestricted use, distribution, and reproduction in any medium, as long as the original authors and source are cited. No permission is required from the authors or the publishers.
Funding Statement
The Tuberculosis and Lung Diseases Research Center, Tabriz University of Medical Sciences, Tabriz, Iran, financially supported the current study.
Introduction
Non-coding RNAs (ncRNAs) constitute the predominant class of transcribed RNAs in eukaryotic cells, and more than 90 % of the entire RNA expression is related to these types of RNAs. These molecules are broadly classified based on length into two major categories: small ncRNAs (sncRNAs, < 200 nt) and long ncRNAs (lncRNAs, > 200 nt).1,2 MicroRNAs (miRNAs), small nucleolar RNAs (snoRNAs), small nuclear RNAs (snRNAs), PIWI-interacting RNA (piRNAs), and small interfering RNAs (siRNAs),3 are the prominent members of sncRNAs.4-6 Conversely, lncRNAs include subtypes such as long intergenic ncRNAs, intronic ncRNAs, macroRNAs, sense ncRNAs, antisense RNAs, and circular RNAs (circRNAs).7-9 In eukaryotic systems, mRNA precursors (pre-mRNAs) commonly contain intronic sequences that are removed via canonical splicing to form mature, linear transcripts.10 However, under certain conditions, these processes can make an entirely different kind of RNA from the same precursor RNA. Initially described approximately 30 years ago, it was discovered that if during non-canonical splicing, specifically back-splicing, an upstream splice acceptor joins a downstream splice donor, circRNAs generating.11,12
This kind of ncRNA contains covalently locked non-stop loop constructions (D-loop) without terminal 5’ caps and 3’ poly-A tails.12,13 It is theoretically possible for all internal exons of genes, excluding the first and last, to give rise to circRNAs. Although back-splicing is considered a relatively rare event, there are more than 200,000 exons in the human genome, and in contrast to the low occurrence of back-splicing, 1,000 unique circRNAs can be found in any given cell type.14 Despite their generally low expression levels, circRNAs exhibit resistance to exonuclease-mediated degradation due to their circular structure and have been implicated in several regulatory roles.15 These include modulation of parental gene expression, alternative splicing or translation, acting as miRNA or RNA-binding protein (RBP) sponges, translation into peptides/ proteins (only a few circRNAs), and the generation of some pseudogenes.16
An increasing number of studies have revealed the aberrant expression of circRNAs in various pathological conditions, including cancers, neurological disorders, and cardiovascular diseases. In oncology, circRNAs can act as either oncogenes or tumor suppressors, depending on their targets and interactions.17 For instance, circHIPK3 promotes colorectal cancer progression by sponging multiple tumor-suppressive miRNAs,18 while circMTO1 suppresses hepatocellular carcinoma (HCC) via inhibition of the oncogenic miR-9. The stability and specific expression patterns of circRNAs in different tissues make them promising candidates for non-invasive diagnostic and prognostic biomarkers, as well as therapeutic targets.19 Accordingly, continued research into the biogenesis, functions, and therapeutic potential of circRNAs is anticipated to yield new insights for scientific exploration and medical innovation. In this review, we summarize the expanding findings on circRNAs and provide an up-to-date account of their biogenesis, regulatory mechanisms, and cellular functions in carcinogenesis.
Biogenesis and functional roles of circRNAs
Biogenesis of circRNAs
In eukaryotic cells, alternative splicing converts pre-RNA into linear mRNA.16 On the other hand, circRNAs are formed through aberrant RNA splicing, specifically back-splicing, which is different from canonical splicing. Approximately 80% of circRNAs are derived from exons, but they can also originate from other parts of the genome, like introns, non-coding regions, antisense strands, and untranslated regions (UTRs).2 Back-splicing generates numerous different circRNAs from a single gene locus, contributing to the complexity of circRNA formation.20 Based on sequence arrangement, circRNAs are classified as exonic circRNAs (ecircRNAs), which contain exon sequences; circular intronic RNAs (ciRNAs), which originate from introns; exonic-intronic circRNAs (EIciRNAs), containing both exonic and intronic sequences; and tRNA intronic circRNAs (tricRNAs), which are formed from spliced tRNA introns.21-23 Although the majority of circRNAs reside in the cytoplasm, EIciRNAs mostly remain in the nucleus.24,25
RNA-binding proteins (RBPs) play a crucial role in the regulation of circRNAs synthesis. RBPs like Quaking (QKI), Muscleblind (MBL/MBNL1), and Fused-in Sarcoma (FUS) can bind to specific motifs on the flanking introns of immature linear RNA.26-28 These RBPs bring the flanking introns together to facilitate the generation of circRNAs.29 Efficient circRNA production requires certain RNA sequence features are needed. For example, exons that can back-splice are often significantly longer up to three times regular exons which is clear in single-exon circRNAs.30 Also, the presence of reverse complementary sequences in flanking intronic regions, like Alu elements, enhances intron pairing and exon circularization. These regions can be either longer or shorter than typical introns.29,31 Inverted tandem repeats in introns also support circRNA formation, with even short repeats around 35 base pairs being sufficient.32 However, these repeats can sometimes make intron base pairing too stable, which makes it less likely for circRNA formation.33,34 As circRNAs mature, introns might not always be removed and can stay between the circularized exons, resulting in a subtype of circRNA known as exonic-intronic circRNAs (EIciRNAs).35
Despite ongoing research, the exact mechanisms of circRNA biogenesis remain unclear. Three models have been proposed: lariat-driven circularization (exon skipping),36 intron pairing-driven circularization,37 and re-splicing-driven circularization,23 each contributing to our understanding of how these unique RNA molecules are formed. Figure 1a schematically illustrates circRNA biogenesis.One common mechanism is lariat-driven circularization, also known as exon-skipping, where partial folding of pre-mRNA brings the upstream donor site (5’ splice site) and the downstream acceptor site (3’ splice site) into proximity. This allows the donor site to attack the receptor site, resulting in the formation of a lariat structure that is subsequently back-spliced to create a new circRNA. This mechanism is notably stimulated by factors such as tumor necrosis factor (TNF)-α and transforming growth factor (TGF)-β in endothelial cells, producing circRNAs alongside linear mRNA which consists of the remaining exons.36,38
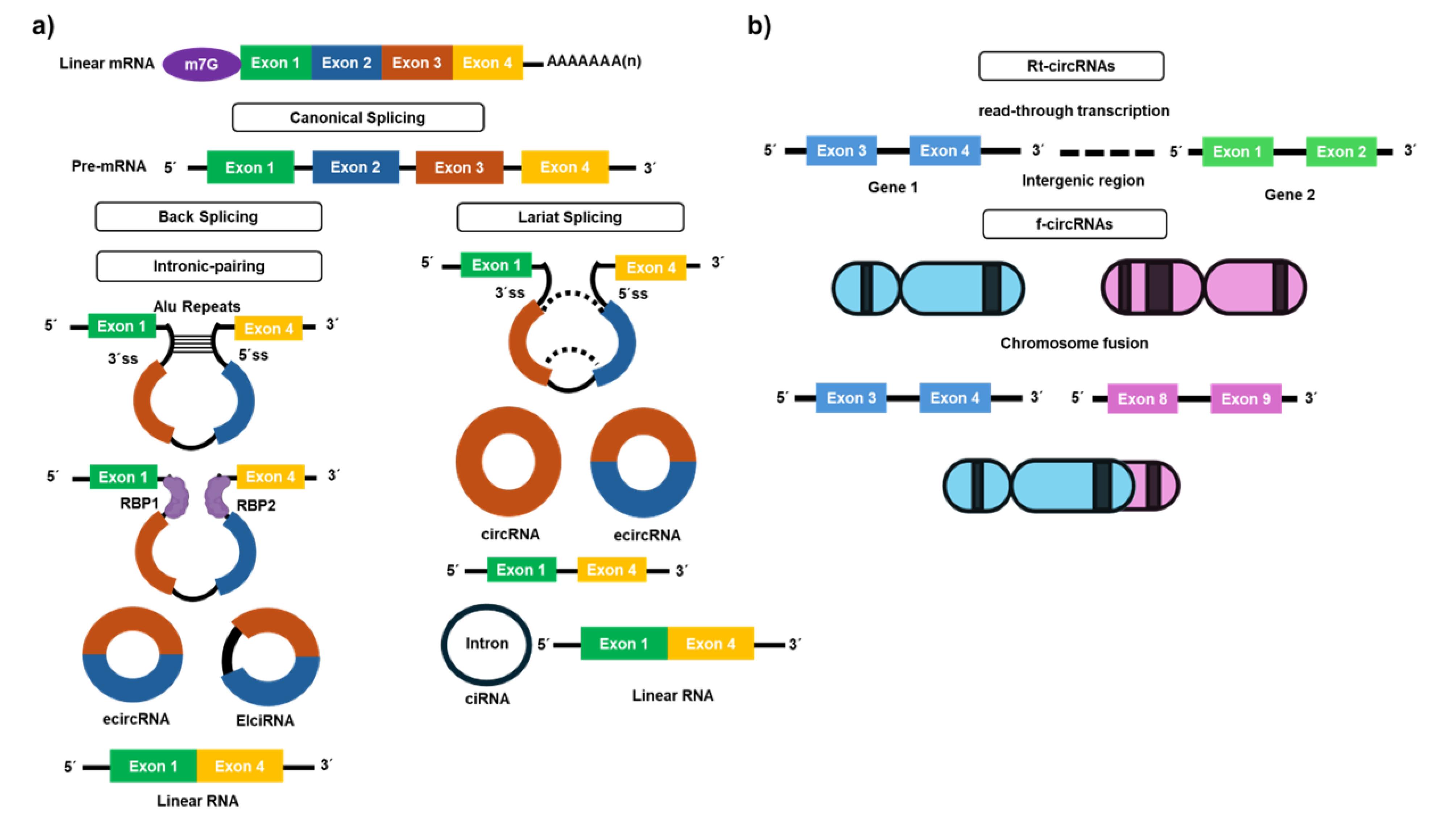
Figure 1.
Biogenesis of circular RNAs and new categories of circular RNAs in oncology. (a) Canonical splicing and Primary methods of back-splicing: intron-pairing promoted by inverted complementary sequences and RNA-binding proteins; lariat creation. (b) Production of rt-circRNAs from read-through transcripts. Generation of f-circRNAs by chromosomal fusions
.
Biogenesis of circular RNAs and new categories of circular RNAs in oncology. (a) Canonical splicing and Primary methods of back-splicing: intron-pairing promoted by inverted complementary sequences and RNA-binding proteins; lariat creation. (b) Production of rt-circRNAs from read-through transcripts. Generation of f-circRNAs by chromosomal fusions
Another key mechanism is intron pairing-driven circularization, which relies on reverse complementary sequences, Alu elements, and the flanking introns. These sequences make possible direct back-splicing. High compatibility between these complementary sequences enhances the circRNA production. This process generates exonic circular RNAs (ecircRNAs) by removing intronic sequences. It also produces exonic-intronic circRNAs (EIciRNAs) that retain some intronic sequences.37,38 A third, lesser-known method is resplicing-driven circularization. Here, a mature linear mRNA undergoes back-splicing to produce circRNAs with one or more exons. The concentration of circRNAs within cells is tightly regulated, with their breakdown being crucial for maintaining cellular function. This degradation process involves the partial activation of endonucleases such as Argonaute 2 (Ago-2), Angiogenin, CPSF73, and RNase L, which create access points for exonucleases to degrade circRNAs completely. Each of these pathways underscores the complex and dynamic nature of circRNA biogenesis and its regulation in cellular biology.23,38,39
Cancer may facilitate the development of novel categories of circRNAs, including read-through circRNAs (rt-circRNAs) and fusion circRNAs (f-circRNAs) (Figure 1b). The rt-circRNAs are derived from read-through transcripts. Read-through transcription occurs when transcription extends over an intergenic region beyond the termination signal, resulting in the synthesis of circRNAs from two neighboring genes. Gene pairs that produce rt-circRNAs are shorter than randomly selected neighboring genes pairs. rt-circRNAs share properties with conventional circRNAs, such as elongated introns and an abundance of repetitive motifs. Read-through circularization may be linked to cancer, characterized by widespread abnormal gene expression mediated by transcription read-through. Of the 460 cancer driver genes, 39 were identified to generate 67 rt-circRNAs, with 31 of them exhibiting cancer-specific expression. Nonetheless, their functional importance in cancer requires further confirmation.40 Cancer-associated chromosomal translocations may result in the generation of fusion-circular RNAs (f-circRNAs). Aberrant chromosomal rearrangements in malignancies may lead to the juxtaposition of two otherwise separated genes, bringing complementary intronic regions into proximity to facilitate reverse splicing. In 2016, Guarnerio et al initially showed that f-circRNAs originate from PML/RARa fusion mRNAs in acute promyelocytic leukemia and that they contribute to carcinogenesis independently of their linear transcripts and protein equivalents, as well as being associated with resistance to anti-cancer therapy. Subsequent investigations have shown that f-circRNAs arise from specific chromosomal translocations, including BCR/ABL1, EML4/ALK, and SLC34A2/ROS1 fusions, seen in both hematological malignancies and solid tumors.41
Mechanisms of action
The ability of circRNAs to control gene expression through diverse mechanisms has led to their increasing recognition as important regulators in cancer biology.42 Their biogenesis often competes with linear mRNA formation, affecting the production of protein. Functionally, circRNAs regulate gene expression via interactions with miRNAs, RNA-binding proteins, and chromatin, and some even assist as templates for translation (Figure 2). In malignancies, their tissue-specific expression, stability, and subcellular localization contribute to their diverse roles, where they influence metastasis, tumorigenesis, and therapy resistance via tumor-suppressive or oncogenic pathways.
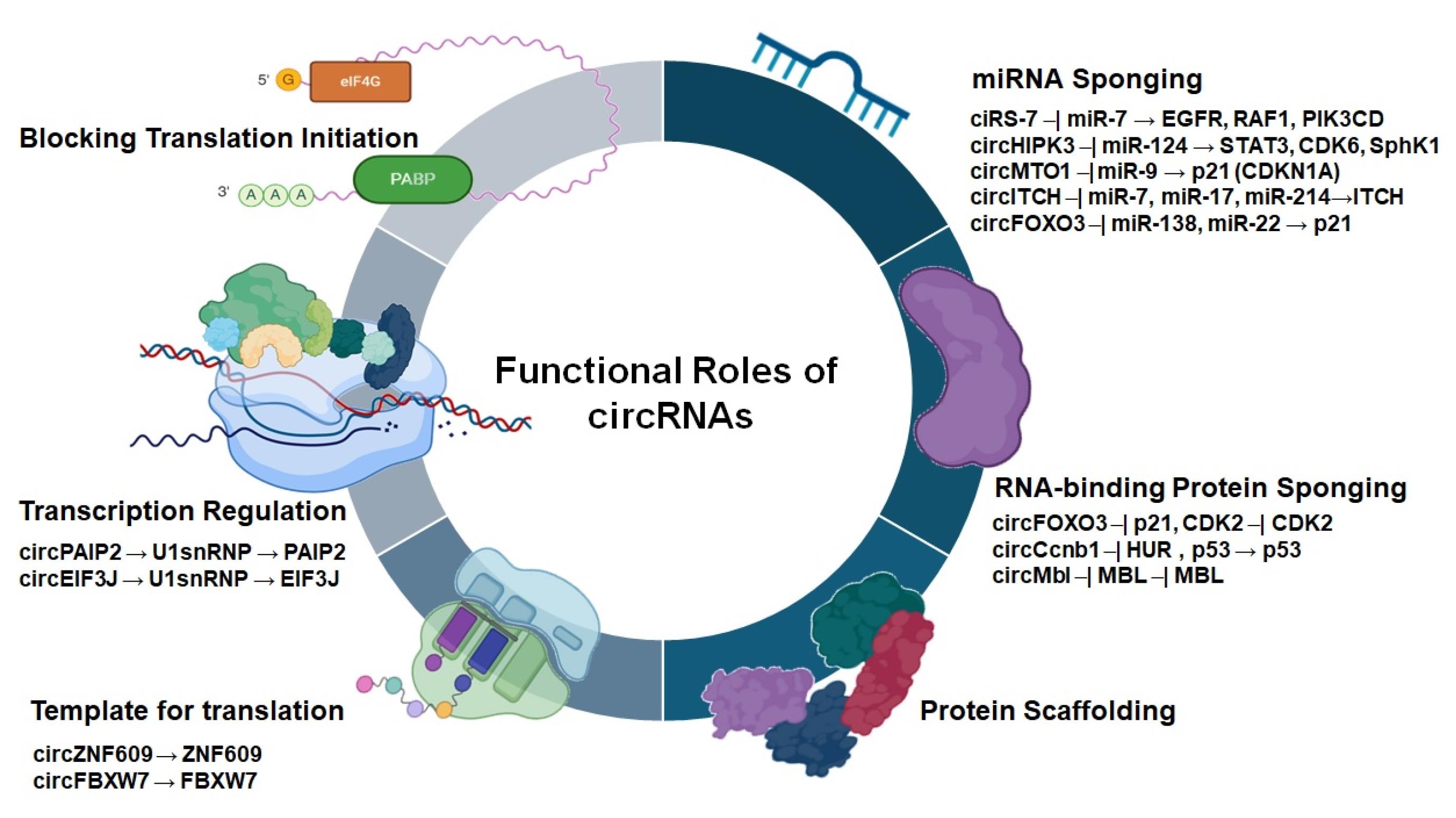
Figure 2.
Schematic representation of known functions of circRNAs
.
Schematic representation of known functions of circRNAs
miRNA sponging
CircRNAs are most known for their function as miRNA sponges, which is among their most extensively studied and well-characterized functions. By containing multiple miRNA response elements (MREs), circRNAs can sequester specific miRNAs and suppress them from repressing their target mRNAs, thus controlling gene expression and protein synthesis. This competitive endogenous RNA (ceRNA) activity has an important role in several physiological and pathological processes, as well as malignancies, osteoarthritis, diabetes, and neurological diseases.43 For instance, ciRS-7 contains over 70 conserved binding sites for miR-7 and has been shown to inhibit its tumor-suppressive activity in several types of malignancy. Similarly, circHIPK3 which is abnormally expressed in cancer tissues, sponges multiple tumor-suppressive miRNAs such as miR-124, miR-193, and miR-637, thus inducing tumor cell invasion, metastasis and development.44 Other circRNAs, as well as circMTO1, circITCH, and circFOXO3, exert tumor-suppressive effects by binding oncogenic miRNAs and regulating key signaling pathways. For example, circMTO1 enhances p21 expression by sponging miR-9 in HCC,19 while circITCH and circFOXO3 target miRNAs such as miR-17, miR-214, miR-224, and miR-9 to prevent tumor development and induce apoptosis. In another study, Circular RNA circ-ABCB10 was shown to induce breast tumor proliferation and development via sponging miR-1271.45 A comparative summary of notable circRNAs, their miRNA targets, related malignancy types, and functional effects is provided in Table 1 to support and contextualize these findings.
Table 1.
CircRNAs as miRNA sponges in various cancers
|
CircRNA
|
Targeted miRNA
|
Functions
|
Cancer type
|
Ref.
|
| ciRS-7 |
miR-7 |
Myocardial infarction; Neural development; anti-oncogenic; stimulates proliferation/metastasis; osteoblastic differentiation insulin secretion |
Various (including breast, liver) |
46
|
| CircHIPK3 |
miR-124, miR-193a, miR-558 |
Stimulates proliferation/migration; prevents cancer progression; β-cell function |
Various (including liver, colorectal) |
47
|
| CircFOXO3 |
miR-138, miR-9, miR-22 |
Cell cycle progression and apoptosis; cardiovascular diseases and cancer |
Various cancers |
48
|
| CircZNF91 |
miR-23b-3p |
Mediates signal transduction between hypoxic and normoxic tumor cells to promote pancreatic cancer chemoresistance |
Pancreatic cancer |
46
|
| CircMTO1 |
miR-9 |
Prevents cancer progression |
HCC |
19
|
| CircCCDC66 |
miR-93, miR-185, miR-33b |
Stimulates cancer progression |
Various (including colorectal) |
49
|
| circIRAK3 |
miR-3607 |
Promotes migration/invasion |
Breast cancer |
50
|
| circRNA_0084043 |
miR-153-3p |
Stimulates cancer progression |
malignant melanoma |
51
|
| CircANKS1B |
miR-148a-3p, miR-152-3p |
Regulation of TGF-β1 signaling pathway |
Breast cancer |
52
|
| Hsa_circ_0008039 |
miR-432-5p |
Increases E2F3 expression |
Breast cancer |
53
|
| circRNA-000911 |
miR-499a |
Regulation of Notch1 and NF-κB signaling pathway |
Breast cancer |
54
|
| CDR1as |
miR-7 |
Prevention of cell proliferation |
Breast, hepatocellular, lung, and gastric cancers |
55,56
|
| circ-ABCB10 |
miR-1271 |
Initiation of cell proliferation |
Breast cancer |
45
|
| circ-ZKSCAN |
N/A |
Prevention of cell proliferation and metastasis |
HCC |
57
|
| circRNA-100269 |
miR-630 |
Prevention of cell proliferation |
HCC |
58
|
| hsa-circ-100338 |
miR-141-3p |
Regulator of metastases |
HCC |
59
|
| hsa_circ_001059 |
miR-30c, miR-122, miR-139-3p, miR-339, miR-1912 |
Regulator for tumor radiotherapy resistance |
Esophageal squamous cell carcinoma |
60
|
| circ-ITCH |
miR-214 |
Prevention of cell proliferation by down-regulation of c-myc, ubiquitination, and degradation of Dvl2 |
Esophageal squamous cell carcinoma, lung cancer, colorectal cancer |
61
|
| circTCF25 |
miR-107, miR-103a-3p |
Initiation of cell proliferation and metastasis |
Bladder cancer |
62
|
| hsa-circ-0043256 |
miR-1252 |
Prevention of cell proliferation |
Lung cancer |
63
|
| Circ-PAX2 |
miR-186 |
Initiation of cell proliferation |
Lung cancer |
63
|
| circEA1 |
miR-372 |
Regulator for cell differentiation and drug resistance |
Lung cancer |
64
|
| Circ-NFIX |
miR-212-3p |
Enhances tumor cell progression |
Lung cancer |
65
|
| hsa-circ-001569 |
miR-145 |
Initiation of cell proliferation and metastasis |
Colorectal cancer |
66
|
| hsa-circ-0000069 |
N/A |
Initiation of cell proliferation and metastasis |
Colorectal cancer |
67
|
| circPVT1 |
miR-125 |
Initiation of cell proliferation and metastasis |
Gastric cancer |
68
|
| circ-LARP4 |
miR-424-5p |
Regulation of tumor progression |
Gastric cancer |
69
|
| circMT01 |
miR-9 |
Regulation of tumor progression |
HCC |
19
|
| hsa_circ_000167 |
miR-181, miR-512, miR-521, miR-556, miR-663 and miR-1204 |
|
Esophageal squamous cell carcinoma |
60
|
| circHIPK3 |
miR-124 |
Regulation of tumor proliferation |
HCC |
70
|
| hsa_circ_0067934 |
miR-98 |
initiation of cell proliferation |
Esophageal squamous cell carcinoma |
71
|
Protein interaction
CircRNAs enable directly interact with RBPs, affecting several cellular processes including cancer development and progression by regulating cellular signaling networks. By harboring RBP binding sites, circRNAs function as molecular scaffolds that facilitate or inhibit protein-protein interactions, control RBPs activity, or prevent them from regulating gene expression. Through the formation of ribonucleoprotein complexes, certain circRNAs, such as ecircRNAs, stabilize these interactions and preserve the functional integrity of the related proteins, thus controlling gene regulation at multiple levels.72 For example, circ-Foxo3 controls cell cycle arrest by interacting with cyclin-dependent kinase 2 (CDK2) and the protein kinase inhibitor p21. In breast tumor, circ-Foxo3 binds to CDK2 and p21, forming a ternary complex that suppress cell cycle proliferation and tumor development.73,74 Additionally, in the context of cellular senescence, circFoxo3 can sequester proteins such as the senescence marker p16 and the transcription factor E2F1, thereby modulating pathways associated with aging and tumor suppression.48 In liver malignancy, circ-Ccnb1 interacts with HuR (ELAVL1) to stabilize CCNB1 mRNA, thus inducing oncogenic cell cycle development.75 Another example, circMbl, forms a binding interaction with the Muscleblind (MBL) protein, which is a pivotal controller of RNA splicing. During this process, circMbl specifically attracts the MBL protein, which plays a crucial role in the alternative splicing of Mbl pre-mRNA.76,77 Furthermore, the decrease of MBL, activation of innate immune dsRNA receptor (PKR) and prevention of Human Antigen R (HuR) protein from binding to Poly(A) Binding Protein Nuclear 1 (PABPN1) mRNA by circPABPN1 can be a result of circRNA activation.78,79 In liver cancer, circ-Ccnb1 sponges miR-194-3p, leading to the promotion of matrix metalloproteinase 9 (MMP-9)-mediated oncogenic effects and inducing tumor progression.80 These RBP-mediated mechanisms underscore the complexity of circRNA functions in cancer, where protein scaffolding and stabilization roles regulate key tumorigenic pathways.81
Transcriptional and translational regulation by circRNAs
In the nucleus, certain circRNAs can to control the gene expression of their host genes. Studies have demonstrated that circRNAs influence the expression of their parental genes through cis-acting mechanisms. In some cases, nuclear circRNAs interact with RNA polymerase II (RNA Pol II) at the promoter region, resulting in the generation of various isoforms of a single gene.82 EiciRNAs are the best-known group of circRNAs with transcriptional activity.83 As an example, circEIF3J and circPAIP2 regulate the Eukaryotic Translation Initiation Factor 3 Subunit J (EIF3J) and Poly(A) Binding Protein Interacting Protein 2 (PAIP2) gene transcriptions by making a complex with the U1 snRNP. This complex then interacts with RNA Pol II, which regulates the transcription of host genes.82,84-86 Circ-ZNF609 and circ-FBXW7 are other examples of transcriptional regulatory circRNAs that are respectively involved in muscular biogenesis and glioma.87,88 The binding of circRNAs to RNA Pol II can control selective splicing by regulation of alternative splice site.
During this process, different splicing sites select pre-mRNAs to produce altered mRNA isoforms. These examples illustrate the diverse roles of circRNAs and highlight their potential as therapeutic targets in cancer and other diseases.89 A new study reveals that circRNAs can compete with their host genes in post-transcriptional processes. Additionally, circRNAs possess internal ribosome entry sites, which enable them to translate independently from the host gene. This model is an intelligent way to regulate stability between the expression levels of circRNAs and host mRNAs.90 As a result, circRNAs control protein production at the transcriptional or post-transcriptional levels. CircZNF609, c-sirt7, and circMbl are three illustrations of circRNAs with coding probability.91-93 In glioblastoma, circFBXW7 is translated into FBXW7-185aa, a peptide that antagonizes c-Myc and prevents tumor cell progression. These findings determine the important role of circRNAs as regulators not only of RNA dynamics but also of protein-coding potential in malignancy.
Modulating immunity and metabolism
Another developing issue is the participation of circRNAs in the immune response. Specific circRNAs can regulate the function of immune cells, thereby impacting the immune system’s ability to react to infections and disorders. This discovery presents new opportunities for investigating circRNAs as potential therapeutic targets for interventions in immune-related diseases.94 CircRNAs influence metabolic pathways. They interact with enzymes and other regulatory factors that control metabolism, which can change how cells process food and use energy. This affects processes such as maintaining blood sugar levels, metabolizing fats, and producing energy. Because circRNAs can do this, they may play a role in health problems related to metabolism, such as obesity and diabetes.95 As we learn more about circRNAs, we can see that these molecules play a key role in how cells control themselves. They can interact with multiple targets within the cell, and they are characterized by high stability and specificity. This property makes circRNAs promising candidates for diagnosing and treating diseases. As we continue to study them and our technology improves, we will learn even more about the function of circRNAs. This will pave the way for novel discoveries and significant advancements in the field of biomedical science.96
Techniques for measuring circRNAs
Measuring and evaluating circRNAs requires special methods because of their unique closed-loop structure, which makes them different from linear RNAs. CircRNA sequencing is a common technique that begins with RNase R treatment to degrade linear RNAs, facilitating the process of circRNA sequencing. This step is crucial as it leaves the circular RNAs intact, allowing for their identification based on unique back-splice junctions through high-depth sequencing, although RNAase treatment is not always mandatory (Figure 3a).22,97-99 CircRNA microarrays offer another high-throughput approach by using probes specifically designed to hybridize with the junction sequences of circRNAs on a solid surface, providing a robust platform for evaluating circRNA expression without requiring RNase treatment, although this can improve accuracy.97,100,101 Northern blotting stands out because it can provide detailed information about the size, isoforms, processing, sequence, and abundance of circRNAs (Figure 3b). It distinguishes between circRNAs and linear RNAs by using different gel electrophoresis methods based on the size of the RNAs.97,102
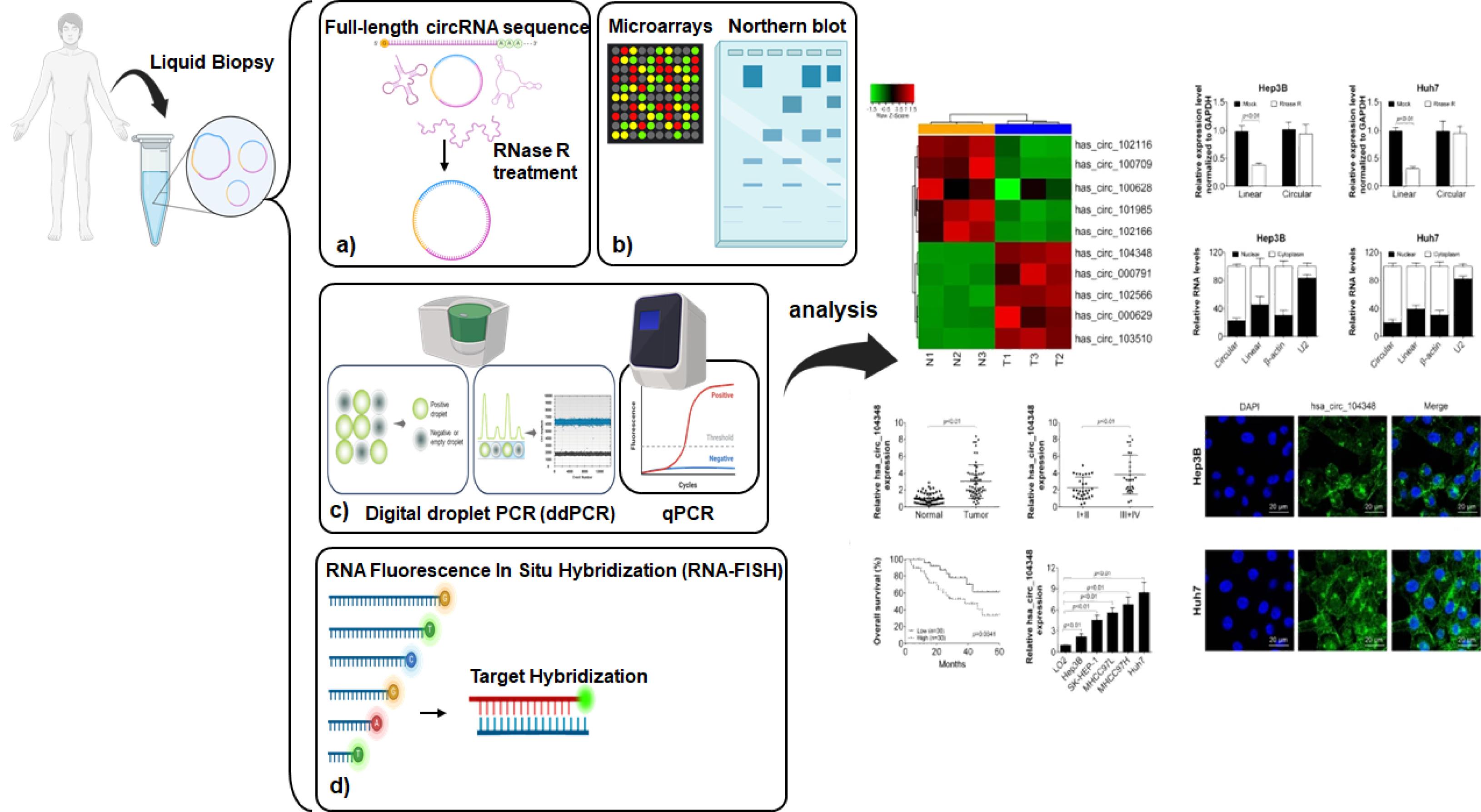
Figure 3.
Schematic representation of diverse circRNA profiling methods
.
Schematic representation of diverse circRNA profiling methods
Real-time quantitative polymerase chain reaction (RT-qPCR) analysis utilizes distinct primers that cover the unique back-splice junctions of circRNAs, enabling accurate quantification while preventing the amplification of linear RNAs. This method can optionally use RNase treatment to increase the concentration of circRNAs, thereby improving measurement accuracy.103 Digital droplet PCR (ddPCR) provides exceptional sensitivity for circRNA quantifying, utilizing nanodroplets for PCR amplification, and determining RNA concentration by comparing the ratio of positive to negative droplets. This method is highly accurate even for low-abundance circRNAs (Figure 3c).104,105
For spatial analysis, RNA Fluorescence in situ Hybridization (RNA-FISH) utilizes probes to detect circRNA junctions. To assess their concentration and distribution inside cells, showing their cellular localization and dynamics (Figure 3d).106 Additionally, to explore how circRNAs interact at the molecular level, methods such as circRNA affinity pulldown, which utilizes biotinylated antisense oligomers (ASOs) to capture circRNAs with streptavidin-coated beads, facilitating interaction mapping within the molecular network. Similarly, immunoprecipitation of circRNA-RBP complexes isolates circRNA-protein complexes using antibodies that target RNA-binding proteins associated with circRNAs. This step allows for later analysis of the RNA component using ddPCR and RT-qPCR. These different methods enhance our understanding of the expression, structure, and functional roles of circRNAs in cellular biology, as well as their potential therapeutic applications.22,97,107
Diagnostic and prognostic potential of circRNAs in cancer
circRNAs have gained attention as important diagnostic and prognostic biomarkers in various malignancies due to their remarkable stability, abundance in body fluids such as plasma and serum, tissue-specific expression pattern, and enabling non-invasive detection (Figure 4).108 Unlike linear RNAs, which typically degrade within 20 hours, the half-life of circRNAs in body fluids generally exceeds 48 hours, which is significantly longer than the half-life of linear RNAs that related to their closed-loop structure and protection within extracellular vesicles. Recent research mention that the half-lives of circRNA vary between 8 and 50 hours, depending on the specific circRNA species and contexts.109 Several researches have shown that expression levels of certain circRNAs are related with clinicopathological characteristics including migration potential, tumor stage, and tumor size, connecting their dysregulation to tumor invasion and development.110 Recent Meta-analyses have demonstrated that circRNAs show good diagnostic performance, with pooled sensitivity and specificity of around 79% and an area under the curve (AUC) of approximately 0.86 in hematological malignancies, demonstrating their potential for early malignancy detection and support clinical management.111 Moreover, combining multiple circRNA into panels considerably improves diagnostic accuracy compared to single circRNAs. For instance, in gastric cancer, a combination of circRNAs improved the AUC from 0.82 to 0.91, representing better sensitivity and specificity.112
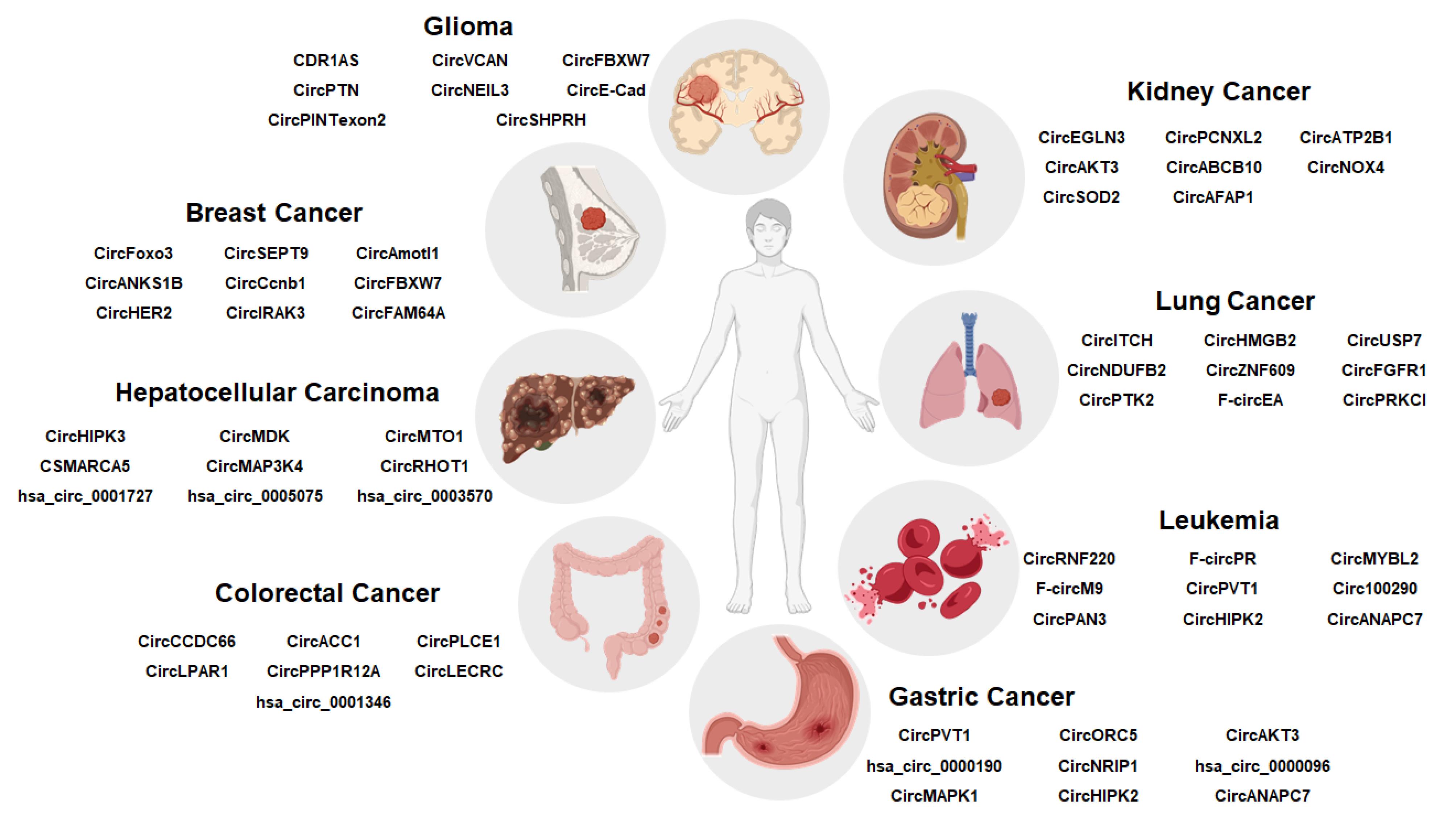
Figure 4.
CircRNAs as biomarkers and therapeutic targets in cancer.This diagram depicts the role of circular RNAs (circRNAs) as biomarkers in cancer diagnosis. The human silhouette in the center represents the patient. Different circRNAs are connected to specific cancer types. These circRNAs enable diagnostic outcomes, such as early detection of tumors, monitoring of tumor progression, and evaluation of treatment responses. The illustration emphasizes the potential of circRNAs as non-invasive biomarkers in liquid biopsy applications for cancer diagnosis
.
CircRNAs as biomarkers and therapeutic targets in cancer.This diagram depicts the role of circular RNAs (circRNAs) as biomarkers in cancer diagnosis. The human silhouette in the center represents the patient. Different circRNAs are connected to specific cancer types. These circRNAs enable diagnostic outcomes, such as early detection of tumors, monitoring of tumor progression, and evaluation of treatment responses. The illustration emphasizes the potential of circRNAs as non-invasive biomarkers in liquid biopsy applications for cancer diagnosis
Similarly, a classifier that employs five circRNAs (circPDLIM5, circSCAF8, circPLXDC2, circSCAMP1, and circCCNT2) extracted from urine extracellular vesicles has shown acceptable performance to recognize high-grade prostate cancer of grade 2 or above.113 In pancreatic ductal adenocarcinoma (PDAC), a panel of five plasma-based liquid biopsy circRNAs (hsa_circ_0060733, hsa_circ_00061117, hsa_circ_0064288, hsa_circ_0007895, and hsa_circ_0007367) was able to distinguish between early stage (stage I/II) and late stage (stage III/IV). Diagnostic accuracy of this circRNA panel for detecting PDAC patients was considerably increased and improved when combined with cancer antigen 19-9 (CA19-9), the conventional biomarker for PDAC.114 On the prognostic side, increased levels of oncogenic circRNAs are associated with poorer overall survival (hazard ratios ranging from 1.3 to 2.3), whereas tumor-suppressive circRNAs are often correlated with better survival rates. These findings underscore circRNAs value in predicting patient prognosis and therapeutic response. Despite these hopeful findings, several challenges and limitation remain, including the lack of standardized detection techniques, large-scale validation studies, and mechanistic insights to fully integrate circRNAs into clinical practice as reliable biomarkers for cancer diagnosis and prognosis are remain.81 hsa-circ-0001649 and hsa-circ-002059 have shown strong potential as biomarkers in human HCC.115,116 Similarly, hsa-circ-001988 has been proposed as a diagnostic marker for gastric cancer.117 While, circPRMT5 was introduced as a biomarker for urothelial carcinoma, where it appears have a role in tumor development and lymph node metastasis.118 Table 2 summarizes circular RNAs that highlights potential biomarkers in several cancer types. As discussed in previous parts of this review, circular RNAs have four recognized functions, and all of them are important in malignancy development.119 Furthermore, circRNAs employ as predictive markers for malignancy treatment efficacy and resistance. Several circRNAs have been recognized for their role in controlling chemosensitivity, underscoring their value in guiding treatment strategies.
Table 2.
CircRNAs with prognostic values in cancers.
|
circRNA
|
Cancer
|
Model Used
|
Mode of Action
|
Translational Stage
|
Related pathological features
|
Ref.
|
| circPRMT5 |
Urothelial carcinoma |
In vitro (UCB cell lines), In vivo (BALB/c nude mice xenograft metastasis model) |
Sponging miR-30c/upregulates SNAIL1/induces/EMTl promotes metastasis |
Preclinical (patient tissue, mouse, exosome-based biomarker) |
Tumor progression and lymph node metastasis |
118
|
| hsa_circ_0001874 |
OSCC |
Human salivary samples (clinical patient cohort, 90 OSCC vs. 82 controls) |
miRNA sponge (targets: miR-661, miR-662, miR-593-5p, miR-107, miR-103a-3p); linked to TNM stage and tumor grade |
Biomarker validation |
Tumor progression, TNM stage, tumor grade |
120
|
| hsa_circ_0001971 |
OSCC |
Human salivary samples (clinical patient cohort, 90 OSCC vs. 82 controls) |
miRNA sponge (targets: miR-152-5p, miR-103a-3p, miR-107, miR-505-3p, miR-9-5p); associated with TNM stage |
Biomarker validation |
Tumor progression, TNM stage, tumor grade |
120
|
| hsa-circ-0001785 |
Breast cancer |
Human plasma samples (n = 57 patients) |
Presumed miRNA sponge (not directly confirmed); associated with tumor burden |
Biomarker validation |
TNM stage, histological grade, distant metastasis, surgery response |
121
|
| Circ-ZEB1.33 |
HCC |
Human HCC tissues, adjacent non-tumorous tissues, HCC cell lines |
miRNA sponge for miR-200a-3p/upregulation of CDK6/ enhanced cell proliferation |
Preclinical functional studies |
Tumor proliferation, upregulated in HCC tissues vs. normal |
122
|
| hsa_circ_0005075 |
HCC |
94 paired HCC and adjacent normal tissues; clinicopathological correlation |
Downregulated in HCC; correlated with tumor size and AFP level (suggestive biomarker role) |
Pre-clinical |
Tumor size, AFP level, Edmondson stage, poor differentiation in HCC |
123
|
| hsa-circ-0001649 |
HCC |
60 paired HCC and adjacent non-tumor liver tissues; cell lines |
Downregulated; ROC analysis suggests potential diagnostic biomarker |
Clinical correlation + biomarker discovery |
Tumor size, TNM stage, AFP levels; diagnostic potential (AUC: 0.834) |
115
|
| hsa_circRNA_100855 |
LSCC |
Human LSCC tissues (n = 52 paired samples), qRT-PCR, microarray |
Not mechanistically tested |
Clinical association study |
Overexpressed in LSCC; correlated with T3–T4 grade, lymph node metastasis, supraglottic site, and advanced stage |
124
|
| hsa_circRNA_104912 |
LSCC |
Human LSCC tissues (n = 52 paired samples), qRT-PCR, microarray |
Not mechanistically tested |
Clinical association study |
Downregulated in LSCC; correlated with T3–T4 grade, lymph node metastasis, poor differentiation, and advanced stage |
124
|
| hsa_circ_0003221 circPTK2 |
Bladder cancer |
40 paired patient tissues and blood samples
- Human cell lines (T24, 5637)
- Nude mice xenograft model |
Promotes proliferation and migration; potentially via miRNA sponge (mechanism not fully defined) |
Preclinical (in vitro & in vivo) |
Overexpressed in poorly differentiated tumors, advanced T stage (II–IV), and N2–N3 lymph node metastasis |
125
|
| hsa-circ-001988 |
Colorectal cancer |
31 paired colorectal tumor and adjacent normal tissues
- Clinical correlation with patient data |
Downregulated in CRC; associated with poor differentiation and perineural invasion; possible biomarker |
Preclinical sample-based study |
Decreased expression in tumors; linked to tumor differentiation, perineural invasion; ROC AUC = 0.788 for diagnosis |
126
|
| hsa_circ_002059 |
Gastric cancer |
Human gastric cancer tissues and plasma |
Downregulate expression; potential biomarker; associated with TNM stage and metastasis |
Preclinical diagnostic biomarker study |
Associated with TNM stage, distal metastasis |
127
|
Abbreviations: OSCC, Oral squamous cell carcinoma; HCC, hepatocellular carcinoma; LSCC, Laryngeal squamous cell carcinoma.
For instance, a specific circRNA expression signature has been used to predict the response to immune checkpoint blockade therapy. The ICBcircSig score was validated based on the weighted expression of circTMTC3 and circFAM117B from melanoma in patients receiving anti-PD-1 or combined anti-CTLA4 and anti-PD-1 therapy. This model, representing that the ICBcircSig score has valuable role in predicting immunotherapy efficacy in melanoma patients.128
A growing number of clinical trials are evaluating the potential of circRNAs as diagnostic and prognostic biomarkers in several malignancies (Table 3). For instance, trial NCT05771337 is recruiting breast tumor patients to validate the clinical efficacy of two plasma-based circRNAs (hsa_circ_0001785 and hsa_circ_100219) for early detection, diagnosis and disease monitoring.129 Additionally, NCT06530082 is evaluating a new dendritic cell vaccine based on a circRNA-derived peptide (circFAM53B-219aa) in individuals with advanced solid tumors (https://www.careacross.com/clinical-trials/trial/NCT06530082). In pancreatic cancer, the CIRCUS trial is exploring circRNA panels for early diagnosis and comparing their efficacy with standard markers, such as CA19-9.114 Furthermore, pilot studies are exploring exosomal circular RNAs in cerebrospinal fluid as potential markers for tracking glioma recurrence.130 Also, preclinical studies are evaluating circPVT1 as a predictor of drug resistance in estrogen receptor alpha-positive (ERα + ) breast cancer.131 Collectively, these researches highlight the growing clinical interest in circRNAs as minimally invasive, stable, sensitive, and specific biomarkers for cancer diagnosis, prognosis, and therapeutic monitoring.
Table 3.
Clinical trials investigating circRNAs as biomarkers in cancers
|
Trial
|
Cancer type(s)
|
circRNA(s) studied
|
Approach/Goal
|
Status
|
Ref.
|
| NCT05771337 |
Breast cancer |
hsa_circ_0001785, hsa_circ_100219 |
Diagnostic/prognostic validation in plasma/serum |
Recruiting |
132
|
| NCT06530082 |
Advanced solid tumors |
circFAM53B-219aa (peptide vaccine) |
Immunotherapy, safety, and efficacy assessment |
Phase I/II, ongoing |
129
|
| CIRCUS Trial |
PDAC |
circPDE8A, circRHOBTB3, panel |
Early detection, comparison with CA19-9 |
Preclinical/Clinical |
114
|
| Glioma (pilot studies) |
Glioma |
circSMARCA5, circHIPK3 (exosomal) |
Recurrence monitoring in CSF |
Preclinical |
130
|
| Drug resistance (preclinical) |
Breast cancer (ERα + ) |
circPVT1 |
Predicting tamoxifen resistance |
Preclinical |
131
|
Abbreviation: PDAC, Pancreatic ductal adenocarcinoma.
Therapeutic applications of circRNAs in cancer treatment
Therapeutic approaches that focus on circRNAs present a hopeful and innovative future in the field of cancer treatment. circRNAs possess distinct characteristics, such as their exceptional stability and specific interactions with miRNAs and proteins, which render them highly suitable for therapeutic intervention.133 To further clarify the translational relevance of circRNAs in cancer, Table 4 provides a comparative overview of well-characterized circRNAs, detailing their associated cancer types, experimental models, modes of action, and current translational status. Here, we examine various strategies that have been developed to utilize circRNAs for cancer treatment.
Table 4.
Comparative summary of promising circRNAs across cancer types: experimental models, molecular mechanisms, and translational relevance
|
circRNA
|
Cancer type(s)
|
Model used
|
Mode of action
|
Translational stage
|
Function
|
Ref.
|
| CircHIPK3 |
Colorectal |
In vitro |
Sponges miR-1207-5p; upregulates FMNL2; promotes proliferation and metastasis |
Preclinical (in vitro, tissue analysis) |
Oncogene |
134
|
| circHIPK3 |
Bladder |
In vitro (T24T, UMUC3); In vivo (xenograft, metastasis model in nude mice) |
Sponges miR-558, downregulating heparanase (HPSE), inhibits MMP-9, VEGF, suppresses angiogenesis and metastasis |
Preclinical (in vitro + xenograft mouse model) |
Tumor suppressor |
135
|
| circMTO1 |
HCC |
In vitro (HepG2, SMMC-7721, QGY-7701, SK-Hep1); In vivo (SMMC-LTNM xenograft in nude mice); Human tissue |
Sponges miR-9; increases p21 expression; tumor suppressor |
Preclinical (in vitro, in vivo, and clinical tissue) |
Tumor suppressor |
19
|
| circITCH |
ESCC |
In vitro (Eca-109, TE-1); In vivo (xenograft nude mouse); Clinical tissues |
circITCH acts as a sponge for miR-7, miR-17, and miR-214. suppression of the Wnt/β-catenin signaling pathway |
Preclinical
(cell lines and mouse xenografts) |
Tumor suppressor |
136
|
| CDR1as (ciRS-7) |
HCC |
In vitro (HepG2, MHCC-97H); In vivo (xenograft mouse); |
circRNA Cdr1as sponges miR-1270, promotes proliferation, migration |
Preclinical (in vitro + in vivo) |
Oncogene |
137
|
| CDR1as (ciRS-7) |
Breast cancer triple-negative breast cancer |
In vitro (MDA-MB-231, BT-549); In vivo (nude mouse tail vein metastasis model) |
Sponges miR-1299, upregulates MMP2 and MMP17,promotes migration, invasion, and metastasis |
Preclinical (in vitro, in vivo, and patient tissues) |
Oncogene |
138
|
| circSMARCA5 |
Prostate cancer |
In vitro (DU145, PC3, LNCaP); In vivo (xenograft and metastasis mouse models); Clinical tissues |
Sponges miR-181b-5p and miR-17-3p upregulate TIMP3, inhibit EMT, proliferation, invasion, metastasis |
Preclinical (in vitro + in vivo + tissue validation) |
tumor suppressor |
139
|
| circFBXW7 |
Glioma |
In vitro (U251, U373); In vivo (xenograft nude mice); Clinical samples |
Encodes FBXW7-185aa protein; promotes c-Myc degradation via USP28 competition |
Preclinical (in vitro, in vivo, and clinical tissue) |
Tumor suppressor |
140
|
| circFOXO3 |
Non-small cell lung cancer |
In vitro |
Sponges miR-155, Upregulates FOXO3, a tumor suppressor, inhibits proliferation and invasion |
Preclinical (in vitro + human tissue) |
Tumor suppressor |
141
|
| circFOXO3 |
Prostate cancer |
In vitro (LNCaP, PC-3, DU145, 22Rv1) |
Sponges miR-29a-3p, upregulates SLC25A15, promotes proliferation, suppresses apoptosis |
Preclinical (in vitro + clinical tissue) |
Oncogene |
142
|
| circRNA_100290 |
Colorectal cancer |
In vitro (HCT116, SW620) |
Sponges miR-516b, upregulating FZD4, activates Wnt/β-catenin signaling, promotes proliferation, invasion, migration |
Preclinical (in vitro + clinical tissue analysis) |
oncogene |
143
|
| circPVT1 |
Gastric cancer |
In vitro (AGS, SGC-7901) |
circPVT1 functions as a competing endogenous RNA (ceRNA) by sponging miR-423-5p |
Preclinical (in vitro) |
Oncogene |
144
|
| circPVT1 |
Osteosarcoma |
In vitro (U2OS, MG63); Clinical tissue |
Sponges miR-205-5p indirectly upregulates c-FLIP, promotes EMT, invasion, and metastasis |
Preclinical (in vitro + human tissue) |
Oncogene |
145
|
| circRNA_0025202 |
Breast |
In vitro (MCF-7, MCF7/TR, T47D); In vivo (xenograft nude mice); Clinical samples |
Sponges miR-182-5p; upregulates FOXO3a; suppresses tumor growth |
Preclinical (in vitro + in vivo + clinical tissues) |
Tumor suppressor in HR + breast cancer |
146
|
| circGFRA1 |
Triple-negative breast cancer |
In vitro (MDA-MB-231, BT549, MDA-MB-468); In vivo (xenograft mouse model); Clinical tissues |
Sponges miR-34a, upregulates GFRA1, promotes proliferation, suppresses apoptosis, linked to poor prognosis |
Preclinical (in vitro + in vivo + patient samples) |
Oncogene |
147
|
| circUHRF1 (hsa_circ_0048677) |
HCC |
In vitro (HCC cell lines + NK-92 cells), In vivo (xenograft in NOD/SCID mice) |
Sponges miR-449c-5p → upregulates TIM-3 on NK cells → → induces NK cell exhaustion |
Preclinical validation and retrospective clinical association with anti-PD1 resistance |
Oncogene |
148
|
| circPRKCI |
Lung adenocarcinoma |
In vitro (LUAD cell lines)/In vivo (xenograft) |
Sponges miR-545 and miR-589, leading to upregulation of E2F7, promoting cell proliferation and tumorigenesis |
Preclinical |
Oncogene |
149
|
| circZNF609 |
Breast cancer |
In vitro (MCF7, MDA-MB-231); In vivo (nude mouse xenograft) |
miRNA sponge (miR-145-5p), upregulates p70S6K1, promotes proliferation, migration, invasion |
Preclinical |
Oncogene |
150
|
Abbreviations: ESCC, Esophageal squamous cell carcinoma; HCC, hepatocellular carcinoma; FMNL2, formin-like 2; CDR1as, Cerebellar degeneration-related protein 1 antisense RNA; ceRNA, endogenous RNA; OSCC, Oral squamous cell carcinoma; LUAD, Lung adenocarcinoma; EMT, Epithelial-mesenchymal transition.
circRNAs targeting strategy
Antisense oligonucleotides (ASOs)
One of the prominent approaches is the use of antisense oligonucleotides (ASOs) specifically degrade oncogenic circRNAs. ASOs are synthetic, short nucleotide sequences that regulate the function of target genes. The FDA has approved two ASO drugs for Duchenne muscular dystrophy (DMD) and spinal muscular atrophy (SMA) treatment.151 circHIPK3, which promotes colorectal and esophageal cancer (EC) by sponging tumor-suppressive microRNAs, can be targeted by ASOs to disrupt its oncogenic activity and restore the normal function of these microRNAs. This targeted degradation can significantly inhibit tumor growth and progression.18,152 Chemical modifications such as phosphorothioate backbones and 2’-O-methylation increase nuclease resistance while simultaneously decrease immunogenicity. Despite these advancements, challenges related to delivery efficiency and tissue specificity remain important problem to clinical translation. Advances in circRNA annotation, modify chemical of ASOs, and improvement of targeted delivery systems may enable for the development of circRNA-directed precision therapeutics that control circRNA activity in a selective and effective way.153 In a pivotal study, Legnini et al confirmed that circZNF609 can be selectively silenced using BSJ-specific ASOs, which led to decrease myoblast proliferation without altering the linear transcript.91
CRISPR/Cas9 gene editing
Advanced gene editing technologies such as CRISPR/Cas9 offer the potential to correct dysfunctional circRNAs implicated in cancer. This approach can be employed to delete or modify oncogenic circRNAs, such as circPRKCI in lung adenocarcinoma, thereby restoring normal cellular functions and inhibiting cancer progression.149 CRISPR/Cas9-mediated knockout of specific circRNAs can provide insights into their roles in malignancy and pave the way for targeted therapies.154 The application of CRISPR/Cas9- to target circRNAs shows hopeful new avenue for cancer treatment, particularly in tumors where specific circRNAs are involved in malignancy development, treatment resistance, or migration potential. Since circRNA expression profiles can vary between different tumor types and even among individual patients, CRISPR/Cas9 technology aligns well with the principles of personalized medicine. By using CRISPR technology to target specific circRNAs correlated with poor prognosis or therapeutic resistance, it may be possible to develop highly specific, patient-centered interventions.155 In related work, CRISPR/Cas9 system has been planned to target the telomerase reverse transcriptase (TERT) promoter or its coding regions. These interventions have effectively prevented TERT transcription and decrease telomerase activity which leading to suppressed cell progression and development and increased apoptosis in malignancy cells.156 As of May 2025, no clinical trials have been published, particularly using CRISPR technology to target circRNAs. However, several preclinical investigations have made considerable progress. Notably, Zhang et al developed an improved CRISPR/Cas13d platform that efficiently degrades circRNAs at the RNA level with higher specificity and efficiency than shRNA methods.157
Delivery strategies
Nanoparticle-based delivery systems
Another promising strategy involves the engineering of circRNA-based nanoparticles for targeted drug delivery. These nanoparticles encapsulate circRNAs, shielding them from nuclease-mediated degradation and enabling their effective delivery into tumor cells. Among the several platforms, lipid nanoparticles (LNPs) has been particularly successful in targeting tumors, types, especially in HCC.158,159 In preclinical studies, LNPs loaded with circRNAs promoted apoptosis, and prevent tumor cell proliferation and progression.160,161 You et al designed magnetically responsive nanoplatforms based PEG-PCL-PEI-C14-coated superparamagnetic iron oxide nanoparticles (SPIONs) for delivering siRNA to target circ_0058051, which led to considerable decrease in circRNA expression and noteworthy tumor inhibition in a HCC model, with no observable off-target toxicity.162 Similarly, Shu et al. used chitosan–epigallocatechin gallate (CS–EGCG) nanoparticles to deliver a circSPIRE1 overexpression plasmid via systemic administration. Their finding showed a significant decrease in lung metastasis in renal cell carcinoma model, by promoting epithelial integrity and repressing angiogenesis.163
Exosome-based delivery systems
Exosome-based delivery systems have garnered important attention as a next generation platform to developing circRNA therapeutics. These nano-sized extracellular vesicles, naturally secreted by cells, can be designed to carry circRNAs directly to tumor cells. These exosomes offer several advantages, as well as their innate ability to pass biological barriers and target specific tissues, such as malignant cells. For instance, circ-0025202 was successfully delivered to breast cancer cells via engineered exosomes. This approach led to considerable decrease in tumor progression and metastasis, indicating the therapeutic potential of exosome-mediated circRNA delivery. This therapeutic strategy utilizes the natural stability and efficient cellular uptake mechanisms of exosomes to promote circRNA delivery while reducing toxicity.164
Synthetic circRNAs and tumor suppressor restoration
Synthetic circRNAs
Synthetic circRNAs can be designed to act as sponges for oncogenic miRNAs, preventing these miRNAs from promoting tumor growth and resistance to apoptosis. For example, synthetic circRNAs can be designed to sequester miR-21-5p, an oncogenic microRNA in cancer cells, thereby preventing it from promoting tumor growth and resistance to apoptosis. These synthetic circRNAs can reduce miRNA availability, thus increasing the tumor-suppressive gene expression like RECK and PDCD4.165,166
Upregulating tumor suppressor circRNAs
CircRNAs like circITCH have confirmed tumor-suppressive function by controlling important oncogenic pathways. In bladder cancer, circ-ITCH prevents tumor growth and development by sponging miR-17/miR-224 and leading to increased expression including p21 and PTEN.167 In colorectal tumor, circITCH suppresses cell progression and growth via sponging miR-7, which results in elevated expression ofITCH, a known negative controller of theWnt/β-cateninsignalingpathway.168,169 Likewise, in gastric cancer, both in vitro and in vivo research have confirmed that cir-ITCH have a tumor suppressor function to suppress tumor carcinogenesis by binding to miR-17 and subsequently downregulating the Wnt/β-catenin pathway.This suppressive impact on tumor progression and growth was decreased when miR-17 was reintroduced.170 Despite these hopeful preclinical results, high expression of tumor suppressor circRNAs for treatment goals have several technical and translational challenges. Improvement invector engineering, nanoparticle-baseddelivery system, and chemical modifications are being developed to overcome these barriers and enable the transition of circRNA-based tumor suppressors into clinical applications.171 Table 5 showstumor suppressor circRNAs and preclinical upregulation strategies.
Table 5.
Tumor suppressor circRNAs and preclinical upregulation strategies
|
circRNA
|
Cancer type
|
Mechanism
|
Therapeutic strategy
|
Effect
|
Clinical/preclinical
|
Ref.
|
| circSMARCA5 |
Glioblastoma |
Sponges oncogenic miRNAs; inhibits angiogenesis |
Plasmid overexpression / synthetic circRNA |
↓ Proliferation, ↓ Migration |
preclinical |
173
|
| circFOXO3 |
NIH3T3 cell, B16 cells (mouse melanoma cell line) |
Binds CDK2/p21; blocks cell cycle progression |
Viral vector overexpression |
↑ Apoptosis, ↓ Tumor growth |
preclinical |
174
|
| circITCH |
Colorectal, bladder |
Sponges miRNAs regulating Wnt pathway |
plasmid-based overexpression, Lentiviral delivery |
↓ Wnt signaling, ↓ Cell proliferation |
preclinical |
168,170
|
Immune modulation
Circular RNAs are emerging as both modulators and potential targets in cancer immunotherapy. Moreover, synthetic circRNAs can be designed to modulate the immune response against cancer. These synthetic circRNAs can act as decoys for immune checkpoint proteins, such as PD-L1, thereby enhancing the body’s immune response against tumors.174 Collectively, these therapeutic strategies underscore the versatile potential of circRNAs in cancer treatment, offering novel avenues to target and overcome the molecular complexities of cancer. Table 6 presents the circRNAs involved in immune modulation and their therapeutic potential.
Table 6.
circRNAs involved in immune modulation and therapeutic potential
|
circRNA
|
Cancer type
|
Immune function
|
Therapeutic strategy
|
In vitro/in vivo
|
Ref.
|
| circUHRF1 |
HCC |
Induces NK cell exhaustion via TIM-3 upregulation |
lentiviral shRNA system + anti-PD1 |
in vitro and in vivo |
149
|
| circ-CPA4 |
NSCLC (A549 and H1299 cell lines; BALB/c nude mice xenografts) |
Promotes PD-L1 expression; immune evasion |
siRNA and shRNA-mediated knockdown |
in vitro and in vivo |
176
|
| circEIF3K |
colorectal cancer (HCT116, SW620, FHC) |
Silencing circEIF3K, up-regulate miR-214, reducing PD-L1 expression |
siRNA and lentiviral shRNA |
in vitro and in vivo |
177
|
circRNAs as immune modulators in the tumor microenvironment
Recent research indicates that certain circRNAs control immune checkpoints, cytokine production, T-cell activity, and immune cell infiltration, thus can contribute to tumor immune evasion and escape. One well-studied example is circUHRF1, which is notably highly expressed in HCC, is secreted via exosomes. This circRNA promotes NK cell exhaustion by increase in T-cell Immunoglobulin and Mucin-domain containing-3 (TIM-3) expression. Functional studies have confirmed that knockdown of circUHRF1 restores NK cell cytotoxicity and enhances the efficacy of anti-programmed cell death protein-1 (PD-1) therapy, underscoring its potential as a therapeutic target.148 Similarly, in non-small cell lung cancer, circ-CPA4 facilitates immune escape by spongingmiR-377, resulting in overexpression of PD-L1 and immune evasion and escape. Knockdown of circ-CPA4, sensitize tumors to immune checkpoint blockade, further highlighting the clinical importance of circRNAs in regulating immune responses.175
circRNAs as biomarkers for immunotherapy response
CircRNAs exhibit exceptional stabilityin blood and exosomes, making them attractive candidates for non-invasive biomarkers to predict immunotherapy response. A compelling example is exosomalcircEIF3K, which is derived from cancer-associated fibroblast induces colorectal cancer growth by the miR-214/PD-L1 signaling pathway.176
Clinical challenges in circRNA-based therapeutic
With pay attention to the acceptable promise of circRNA-based therapeutics, the application of circRNAs stay in preclinical phase, introducing these challenges is important for proceeding their development from preclinical studies to clinical applications. This section highlights the important limits of these techniques and explores potential strategies for overcoming them. One important dis-advantage is the risk of off-target gene silencing, where RNA interference (RNAi) techniques including the use of small interfering RNAs (siRNAs), can knockdown unintended genes due to partial complementarity leading to unexpected and mostly damaging effects. Although recent techniques, like CRISPR/Cas13 technology, show higher specificity in targeting circRNAs, but these method and technology still require validation in vivo application, before they can be safely used in clinic.177 Another considerable challenge is related the non-specific delivery of therapeutic agents to tissues or cell types since some circRNAs are expressed in multiple tissue types, this can lead to off-target effects in non-diseased tissues. For this reason, researchers are designing nanoparticle delivery platform that induce the selectivity and accuracy of delivering therapeutic agents to specific tissues or cell.178 Furthermore, numerous technical and safety problems prevent the clinical translation of circRNA-based therapies. For instance, gold nanoparticles (AuNPs), widely used to deliver vehicles for circRNA-targeting agents in vivo models whereas increase toxicity and safety risks.179 Ongoing research is focused on optimizing AuNP properties for improve safety and biocompatibility or selecting safer alternatives, including lipid nanoparticle (LNP)-based systems, which are already approved for use in mRNA-based vaccines and RNA therapies.178 However, the using LNPs also has some disadvantages, including inefficient endosomal escape and limited ability to target solid tumors is the main reason to prevent their application in malignancy treatment therapy.180
In clinical scale, another important disadvantage is the complexity and cost of producing high-purity circRNAs. High expression vectors frequently generate linear or mis-spliced byproducts, decreasing therapeutic purity. Recent studies in template-based in vitro circularization and purification are hopeful but they are not yet suitable for industrial production.181 Moreover, synthetic circRNAs may induce immune responses because they do not have specific post-transcriptional modifications present in endogenous circRNAs, including N6-methyladenosine (m6 A), which help them to evade immune responses. There are some techniques to decrease synthetic circRNAs immunogenicity including chemical modifications and coating synthetic circRNAs with RBPs to escape immune responses.182 From a diagnostic feature, the remarkable stability of circRNAs in body fluids like plasma, serum, and saliva supports their potential application in liquid biopsy platforms. However, the clinical applications of circRNAs stay limited due to the absence of standardized detection protocols and unpredictable functional validation in different patient cohorts. Moreover, ongoing discussion about the coding potential of some circRNAs, emphasizes the need for more complete functional characterization and mechanistic studies.183 Recent studies have also underscored translational setbacks. For instance, overexpression of circRNAs leads to induction of immune responses or inability to reproduce in vitro effects in vivo. to address this challenge, researchers are engineering synthetic circRNAs that more closely mimic endogenous molecules, with the goal of maintaining function and reducing immunogenicity. Additionally, combination therapeutic methods, including pairing circRNA delivery with immune checkpoint inhibitors, are also being investigated to increase efficiency.184 Overcoming these challenges is essential to translating the preclinical findings into effective clinical therapies.
Conclusion
circRNAs are gaining recognitionas important regulators in cancer biology, with diverseroles as diagnostic biomarkers, therapeutic targets, and even direct treatment approaches. Their unique characteristics, such as remarkable stability, functional versatility, and precise regulatory capabilities, make them a promising class of molecules for personalized cancer treatment. Nevertheless, the field remains in its early stages, and inconsistencies in studies, along with technical limitations, underscore the urgent requirement for rigorous and standardized research frameworks. The future clinical impact of circRNAs is dependent on ongoing methodological advancements, scalable delivery systems, and robust clinical validation. As research into circRNAs continues to gain momentum, several remarkable methods and techniques are emerging that could reshape malignancy diagnosis and treatment. The advancement of CRISPR-based technologies and the development of chemically stabilized synthetic circRNAs may improve specificity and durability. Alongside, progress in nanoparticle and exosome-mediated delivery platform provides more targeted and efficient delivery of circRNA-based therapeutics, increasing their clinical potential.185 Regardless of these developments, important challenges remain, such as lack of clinical trial data, functional diversity of circRNAs across various cancer types, and an incomplete understanding of circRNA-host gene interactions. Future studies efforts should prioritize large-scale, standardized investigations accompanied by robust functional validation across a variety ofpreclinical and clinical models. Also, integrating circRNA signatures with genomic and proteomic profiling could facilitate highly personalized cancer management, paving the way for accurate oncology tools that using circRNA biology for personalized prognosis, diagnosis, and treatment interventions.81,186
Competing Interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Consent for Publication
Not applicable.
Data Availability Statement
Not applicable.
Ethical Approval
Not applicable.
Acknowledgements
We would like to appreciate the cooperation of the Clinical Research Development Unit, Imam Reza General Hospital, Tabriz, Iran, in conducting this research.
References
- Beňačka R, Szabóová D, Guľašová Z, Hertelyová Z, Radoňak J. Non-coding RNAs in human cancer and other diseases: overview of the diagnostic potential. Int J Mol Sci 2023; 24(22):16213. doi: 10.3390/ijms242216213 [Crossref] [ Google Scholar]
- Aftabi Y, Ansarin K, Shanehbandi D, Khalili M, Seyedrezazadeh E, Rahbarnia L. Long non-coding RNAs as potential biomarkers in the prognosis and diagnosis of lung cancer: a review and target analysis. IUBMB Life 2021; 73(2):307-27. doi: 10.1002/iub.2430 [Crossref] [ Google Scholar]
- Mansoori B, Sandoghchian Shotorbani S, Baradaran B. RNA interference and its role in cancer therapy. Adv Pharm Bull 2014; 4(4):313-21. doi: 10.5681/apb.2014.046 [Crossref] [ Google Scholar]
- Esteller M. Non-coding RNAs in human disease. Nat Rev Genet 2011; 12(12):861-74. doi: 10.1038/nrg3074 [Crossref] [ Google Scholar]
- Shanehbandi D, Asadi M, Seyedrezazadeh E, Zafari V, Shekari N, Akbari M. MicroRNA-based biomarkers in lung cancer: recent advances and potential applications. Curr Mol Med 2023; 23(7):648-67. doi: 10.2174/2772432817666220520085719 [Crossref] [ Google Scholar]
- Kamran S, Seyedrezazadeh E, Shanehbandi D, Asadi M, Zafari V, Shekari N. Combination therapy with KRAS and P38α siRNA suppresses colorectal cancer growth and development in SW480 cell line. J Gastrointest Cancer 2022; 53(3):597-604. doi: 10.1007/s12029-021-00667-1 [Crossref] [ Google Scholar]
- Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet 2009; 10(3):155-9. doi: 10.1038/nrg2521 [Crossref] [ Google Scholar]
- Romano G, Veneziano D, Acunzo M, Croce CM. Small non-coding RNA and cancer. Carcinogenesis 2017; 38(5):485-91. doi: 10.1093/carcin/bgx026 [Crossref] [ Google Scholar]
- Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao L, Zhang Y, et al. The landscape of circular RNA in cancer. Cell 2019;176(4):869-81.e13. doi: 10.1016/j.cell.2018.12.021.
- Will CL, Lührmann R. Spliceosome structure and function. Cold Spring HarbPerspect Biol 2011; 3(7):a003707. doi: 10.1101/cshperspect.a003707 [Crossref] [ Google Scholar]
- Ivanov A, Memczak S, Wyler E, Torti F, Porath HT, Orejuela MR. Analysis of intron sequences reveals hallmarks of circular RNA biogenesis in animals. Cell Rep 2015; 10(2):170-7. doi: 10.1016/j.celrep.2014.12.019 [Crossref] [ Google Scholar]
- Wang J, Wang L. Deep learning of the back-splicing code for circular RNA formation. Bioinformatics 2019; 35(24):5235-42. doi: 10.1093/bioinformatics/btz382 [Crossref] [ Google Scholar]
- Xiao MS, Ai Y, Wilusz JE. Biogenesis and functions of circular RNAs come into focus. Trends Cell Biol 2020; 30(3):226-40. doi: 10.1016/j.tcb.2019.12.004 [Crossref] [ Google Scholar]
- Zhang XO, Dong R, Zhang Y, Zhang JL, Luo Z, Zhang J. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res 2016; 26(9):1277-87. doi: 10.1101/gr.202895.115 [Crossref] [ Google Scholar]
- Ebbesen KK, Hansen TB, Kjems J. Insights into circular RNA biology. RNA Biol 2017; 14(8):1035-45. doi: 10.1080/15476286.2016.1271524 [Crossref] [ Google Scholar]
- Hsiao KY, Sun HS, Tsai SJ. Circular RNA - new member of noncoding RNA with novel functions. Exp Biol Med (Maywood) 2017; 242(11):1136-41. doi: 10.1177/1535370217708978 [Crossref] [ Google Scholar]
- Taheri M, Najafi S, Basiri A, Hussen BM, Baniahmad A, Jamali E. The role and clinical potentials of circular RNAs in prostate cancer. Front Oncol 2021; 11:781414. doi: 10.3389/fonc.2021.781414 [Crossref] [ Google Scholar]
- Zeng K, Chen X, Xu M, Liu X, Hu X, Xu T. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis 2018; 9(4):417. doi: 10.1038/s41419-018-0454-8 [Crossref] [ Google Scholar]
- Han D, Li J, Wang H, Su X, Hou J, Gu Y. Circular RNA circMTO1 acts as the sponge of microRNA-9 to suppress hepatocellular carcinoma progression. Hepatology 2017; 66(4):1151-64. doi: 10.1002/hep.29270 [Crossref] [ Google Scholar]
- Araghi M, Soerjomataram I, Jenkins M, Brierley J, Morris E, Bray F. Global trends in colorectal cancer mortality: projections to the year 2035. Int J Cancer 2019; 144(12):2992-3000. doi: 10.1002/ijc.32055 [Crossref] [ Google Scholar]
- Lasda E, Parker R. Circular RNAs: diversity of form and function. RNA 2014; 20(12):1829-42. doi: 10.1261/rna.047126.114 [Crossref] [ Google Scholar]
- Jeck WR, Sharpless NE. Detecting and characterizing circular RNAs. Nat Biotechnol 2014; 32(5):453-61. doi: 10.1038/nbt.2890 [Crossref] [ Google Scholar]
- Chen I, Chen CY, Chuang TJ. Biogenesis, identification, and function of exonic circular RNAs. Wiley Interdiscip Rev RNA 2015; 6(5):563-79. doi: 10.1002/wrna.1294 [Crossref] [ Google Scholar]
- Greene J, Baird AM, Brady L, Lim M, Gray SG, McDermott R. Circular RNAs: biogenesis, function and role in human diseases. Front Mol Biosci 2017; 4:38. doi: 10.3389/fmolb.2017.00038 [Crossref] [ Google Scholar]
- Ebbesen KK, Kjems J, Hansen TB. Circular RNAs: identification, biogenesis and function. BiochimBiophys Acta 2016; 1859(1):163-8. doi: 10.1016/j.bbagrm.2015.07.007 [Crossref] [ Google Scholar]
- Patop IL, Wüst S, Kadener S. Past, present, and future of circRNAs. EMBO J 2019; 38(16):e100836. doi: 10.15252/embj.2018100836 [Crossref] [ Google Scholar]
- Li M, Ding W, Sun T, Tariq MA, Xu T, Li P. Biogenesis of circular RNAs and their roles in cardiovascular development and pathology. FEBS J 2018; 285(2):220-32. doi: 10.1111/febs.14191 [Crossref] [ Google Scholar]
- Pandey PR, Rout PK, Das A, Gorospe M, Panda AC. RPAD (RNase R treatment, polyadenylation, and poly(A) + RNA depletion) method to isolate highly pure circular RNA. Methods 2019; 155:41-8. doi: 10.1016/j.ymeth.2018.10.022 [Crossref] [ Google Scholar]
- Ashwal-Fluss R, Meyer M, Pamudurti NR, Ivanov A, Bartok O, Hanan M. circRNA biogenesis competes with pre-mRNA splicing. Mol Cell 2014; 56(1):55-66. doi: 10.1016/j.molcel.2014.08.019 [Crossref] [ Google Scholar]
- Cao D. Reverse complementary matches simultaneously promote both back-splicing and exon-skipping. BMC Genomics 2021; 22(1):586. doi: 10.1186/s12864-021-07910-w [Crossref] [ Google Scholar]
- Daniel C, Behm M, Öhman M. The role of Alu elements in the cis-regulation of RNA processing. Cell Mol Life Sci 2015; 72(21):4063-76. doi: 10.1007/s00018-015-1990-3 [Crossref] [ Google Scholar]
- Barrett SP, Wang PL, Salzman J. Circular RNA biogenesis can proceed through an exon-containing lariat precursor. Elife 2015; 4:e07540. doi: 10.7554/eLife.07540 [Crossref] [ Google Scholar]
- Bachvaroff TR, Place AR. From stop to start: tandem gene arrangement, copy number and trans-splicing sites in the dinoflagellate Amphidiniumcarterae. PLoS One 2008; 3(8):e2929. doi: 10.1371/journal.pone.0002929 [Crossref] [ Google Scholar]
- Aravin AA, Naumova NM, Tulin AV, Vagin VV, Rozovsky YM, Gvozdev VA. Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D melanogaster germline. Curr Biol 2001; 11(13):1017-27. doi: 10.1016/s0960-9822(01)00299-8 [Crossref] [ Google Scholar]
- Shi Y, Fang N, Li Y, Guo Z, Jiang W, He Y. Circular RNA LPAR3 sponges microRNA-198 to facilitate esophageal cancer migration, invasion, and metastasis. Cancer Sci 2020; 111(8):2824-36. doi: 10.1111/cas.14511 [Crossref] [ Google Scholar]
- Yin Y, Long J, He Q, Li Y, Liao Y, He P. Emerging roles of circRNA in formation and progression of cancer. J Cancer 2019; 10(21):5015-21. doi: 10.7150/jca.30828 [Crossref] [ Google Scholar]
- Yao T, Chen Q, Fu L, Guo J. Circular RNAs: biogenesis, properties, roles, and their relationships with liver diseases. Hepatol Res 2017; 47(6):497-504. doi: 10.1111/hepr.12871 [Crossref] [ Google Scholar]
- Chen LL, Yang L. Regulation of circRNA biogenesis. RNA Biol 2015; 12(4):381-8. doi: 10.1080/15476286.2015.1020271 [Crossref] [ Google Scholar]
- Ragan C, Goodall GJ, Shirokikh NE, Preiss T. Insights into the biogenesis and potential functions of exonic circular RNA. Sci Rep 2019; 9(1):2048. doi: 10.1038/s41598-018-37037-0 [Crossref] [ Google Scholar]
- Vidal AF. Read-through circular RNAs reveal the plasticity of RNA processing mechanisms in human cells. RNA Biol 2020; 17(12):1823-6. doi: 10.1080/15476286.2020.1805233 [Crossref] [ Google Scholar]
- Guarnerio J, Bezzi M, Jeong JC, Paffenholz SV, Berry K, Naldini MM. Oncogenic role of fusion-circRNAs derived from cancer-associated chromosomal translocations. Cell 2016; 165(2):289-302. doi: 10.1016/j.cell.2016.03.020 [Crossref] [ Google Scholar]
- Wu X, Tao P, Zhou Q, Li J, Yu Z, Wang X. IL-6 secreted by cancer-associated fibroblasts promotes epithelial-mesenchymal transition and metastasis of gastric cancer via JAK2/STAT3 signaling pathway. Oncotarget 2017; 8(13):20741-50. doi: 10.18632/oncotarget.15119 [Crossref] [ Google Scholar]
- Dori M, Bicciato S. Integration of bioinformatic predictions and experimental data to identify circRNA-miRNA associations. Genes (Basel) 2019; 10(9):642. doi: 10.3390/genes10090642 [Crossref] [ Google Scholar]
- Wen Y, Li B, He M, Teng S, Sun Y, Wang G. circHIPK3 promotes proliferation and migration and invasion via regulation of miR-637/HDAC4 signaling in osteosarcoma cells. Oncol Rep 2021; 45(1):169-79. doi: 10.3892/or.2020.7833 [Crossref] [ Google Scholar]
- Liang HF, Zhang XZ, Liu BG, Jia GT, Li WL. Circular RNA circ-ABCB10 promotes breast cancer proliferation and progression through sponging miR-1271. Am J Cancer Res 2017; 7(7):1566-76. [ Google Scholar]
- Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK. Natural RNA circles function as efficient microRNA sponges. Nature 2013; 495(7441):384-8. doi: 10.1038/nature11993 [Crossref] [ Google Scholar]
- Gu A, Jaijyan DK, Yang S, Zeng M, Pei S, Zhu H. Functions of circular RNA in human diseases and illnesses. Noncoding RNA 2023; 9(4):38. doi: 10.3390/ncrna9040038 [Crossref] [ Google Scholar]
- Du WW, Yang W, Chen Y, Wu ZK, Foster FS, Yang Z. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur Heart J 2017; 38(18):1402-12. doi: 10.1093/eurheartj/ehw001 [Crossref] [ Google Scholar]
- Hsiao KY, Lin YC, Gupta SK, Chang N, Yen L, Sun HS. Noncoding effects of circular RNA CCDC66 promote colon cancer growth and metastasis. Cancer Res 2017; 77(9):2339-50. doi: 10.1158/0008-5472.Can-16-1883 [Crossref] [ Google Scholar]
- Wu J, Jiang Z, Chen C, Hu Q, Fu Z, Chen J. CircIRAK3 sponges miR-3607 to facilitate breast cancer metastasis. Cancer Lett 2018; 430:179-92. doi: 10.1016/j.canlet.2018.05.033 [Crossref] [ Google Scholar]
- Luan W, Shi Y, Zhou Z, Xia Y, Wang J. circRNA_0084043 promote malignant melanoma progression via miR-153-3p/Snail axis. BiochemBiophys Res Commun 2018; 502(1):22-9. doi: 10.1016/j.bbrc.2018.05.114 [Crossref] [ Google Scholar]
- Zeng K, He B, Yang BB, Xu T, Chen X, Xu M. The pro-metastasis effect of circANKS1B in breast cancer. Mol Cancer 2018; 17(1):160. doi: 10.1186/s12943-018-0914-x [Crossref] [ Google Scholar]
- Liu Y, Lu C, Zhou Y, Zhang Z, Sun L. Circular RNA hsa_circ_0008039 promotes breast cancer cell proliferation and migration by regulating miR-432-5p/E2F3 axis. BiochemBiophys Res Commun 2018; 502(3):358-63. doi: 10.1016/j.bbrc.2018.05.166 [Crossref] [ Google Scholar]
- Wang H, Xiao Y, Wu L, Ma D. Comprehensive circular RNA profiling reveals the regulatory role of the circRNA-000911/miR-449a pathway in breast carcinogenesis. Int J Oncol 2018; 52(3):743-54. doi: 10.3892/ijo.2018.4265 [Crossref] [ Google Scholar]
- Zhong Q, Huang J, Wei J, Wu R. Circular RNA CDR1as sponges miR-7-5p to enhance E2F3 stability and promote the growth of nasopharyngeal carcinoma. Cancer Cell Int 2019; 19:252. doi: 10.1186/s12935-019-0959-y [Crossref] [ Google Scholar]
- Hansen TB, Kjems J, Damgaard CK. Circular RNA and miR-7 in cancer. Cancer Res 2013; 73(18):5609-12. doi: 10.1158/0008-5472.Can-13-1568 [Crossref] [ Google Scholar]
- Yao Z, Luo J, Hu K, Lin J, Huang H, Wang Q. ZKSCAN1 gene and its related circular RNA (circZKSCAN1) both inhibit hepatocellular carcinoma cell growth, migration, and invasion but through different signaling pathways. Mol Oncol 2017; 11(4):422-37. doi: 10.1002/1878-0261.12045 [Crossref] [ Google Scholar]
- Lei B, Tian Z, Fan W, Ni B. Circular RNA: a novel biomarker and therapeutic target for human cancers. Int J Med Sci 2019; 16(2):292-301. doi: 10.7150/ijms.28047 [Crossref] [ Google Scholar]
- Huang XY, Huang ZL, Xu YH, Zheng Q, Chen Z, Song W. Comprehensive circular RNA profiling reveals the regulatory role of the circRNA-100338/miR-141-3p pathway in hepatitis B-related hepatocellular carcinoma. Sci Rep 2017; 7(1):5428. doi: 10.1038/s41598-017-05432-8 [Crossref] [ Google Scholar]
- Han C, Seebacher NA, Hornicek FJ, Kan Q, Duan Z. Regulation of microRNAs function by circular RNAs in human cancer. Oncotarget 2017; 8(38):64622-37. doi: 10.18632/oncotarget.19930 [Crossref] [ Google Scholar]
- Wan L, Zhang L, Fan K, Cheng ZX, Sun QC, Wang JJ. Circular RNA-ITCH suppresses lung cancer proliferation via inhibiting the Wnt/β-catenin pathway. Biomed Res Int 2016; 2016:1579490. doi: 10.1155/2016/1579490 [Crossref] [ Google Scholar]
- Cong L, Yang Q, Hu C, Yu Q, Hao S, Li D. Current status of functional studies on circular RNAs in bladder cancer and their potential role as diagnostic and prognostic biomarkers: a review. Med Sci Monit 2019; 25:3425-34. doi: 10.12659/msm.916697 [Crossref] [ Google Scholar]
- Wang L, Xu C, Wang C, Gong W, Zhang K, Chen Q. Circ-PAX2 promotes proliferation and metastasis by absorbing miR-186 in lung cancer cells. Int J Clin Exp Pathol 2018; 11(7):3793-801. [ Google Scholar]
- Lizarbe MA, Calle-Espinosa J, Fernández-Lizarbe E, Fernández-Lizarbe S, Robles M, Olmo N. Colorectal cancer: from the genetic model to posttranscriptional regulation by noncoding RNAs. Biomed Res Int 2017; 2017:7354260. doi: 10.1155/2017/7354260 [Crossref] [ Google Scholar]
- Lu J, Zhu Y, Qin Y, Chen Y. CircNFIX acts as a miR-212-3p sponge to enhance the malignant progression of non-small cell lung cancer by up-regulating ADAM10. Cancer Manag Res 2020; 12:9577-87. doi: 10.2147/cmar.S272309 [Crossref] [ Google Scholar]
- Xie H, Ren X, Xin S, Lan X, Lu G, Lin Y. Emerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancer. Oncotarget 2016; 7(18):26680-91. doi: 10.18632/oncotarget.8589 [Crossref] [ Google Scholar]
- Wang HY, Wang YP, Zeng X, Zheng Y, Guo QH, Ji R. Circular RNA is a popular molecule in tumors of the digestive system (Review). Int J Oncol 2020; 57(1):21-42. doi: 10.3892/ijo.2020.5054 [Crossref] [ Google Scholar]
- Kun-Peng Z, Xiao-Long M, Chun-Lin Z. Overexpressed circPVT1, a potential new circular RNA biomarker, contributes to doxorubicin and cisplatin resistance of osteosarcoma cells by regulating ABCB1. Int J Biol Sci 2018; 14(3):321-30. doi: 10.7150/ijbs.24360 [Crossref] [ Google Scholar]
- Zhang J, Liu H, Hou L, Wang G, Zhang R, Huang Y. Circular RNA_LARP4 inhibits cell proliferation and invasion of gastric cancer by sponging miR-424-5p and regulating LATS1 expression. Mol Cancer 2017; 16(1):151. doi: 10.1186/s12943-017-0719-3 [Crossref] [ Google Scholar]
- Chen G, Shi Y, Liu M, Sun J. circHIPK3 regulates cell proliferation and migration by sponging miR-124 and regulating AQP3 expression in hepatocellular carcinoma. Cell Death Dis 2018; 9(2):175. doi: 10.1038/s41419-017-0204-3 [Crossref] [ Google Scholar]
- Xia W, Qiu M, Chen R, Wang S, Leng X, Wang J. Circular RNA has_circ_0067934 is upregulated in esophageal squamous cell carcinoma and promoted proliferation. Sci Rep 2016; 6:35576. doi: 10.1038/srep35576 [Crossref] [ Google Scholar]
- Qu S, Liu Z, Yang X, Zhou J, Yu H, Zhang R. The emerging functions and roles of circular RNAs in cancer. Cancer Lett 2018; 414:301-9. doi: 10.1016/j.canlet.2017.11.022 [Crossref] [ Google Scholar]
- Lu WY. Roles of the circular RNA circ-Foxo3 in breast cancer progression. Cell Cycle 2017; 16(7):589-90. doi: 10.1080/15384101.2017.1278935 [Crossref] [ Google Scholar]
- Du WW, Fang L, Yang W, Wu N, Awan FM, Yang Z. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ 2017; 24(2):357-70. doi: 10.1038/cdd.2016.133 [Crossref] [ Google Scholar]
- Rong MH, Li JD, Zhong LY, Huang YZ, Chen J, Xie LY. CCNB1 promotes the development of hepatocellular carcinoma by mediating DNA replication in the cell cycle. Exp Biol Med (Maywood) 2022; 247(5):395-408. doi: 10.1177/15353702211049149 [Crossref] [ Google Scholar]
- Pamudurti NR, Konakondla-Jacob VV, Krishnamoorthy A, Ashwal-Fluss R, Bartok O, Wüst S, et al. An in vivo knockdown strategy reveals multiple functions for circMbl. bioRxiv [Preprint]. November 29, 2018. Available from: https://www.biorxiv.org/content/10.1101/483271v1.
- Konakondla JV. Molecular Interaction of circMbl and MBL in Vivo [dissertation]. Brandeis University Graduate School of Arts and Sciences; 2019.
- Li XX, Xiao L, Chung HK, Ma XX, Liu X, Song JL. Interaction between HuR and circPABPN1 modulates autophagy in the intestinal epithelium by altering ATG16L1 translation. Mol Cell Biol 2020; 40(6):e00492-19. doi: 10.1128/mcb.00492-19 [Crossref] [ Google Scholar]
- Abdelmohsen K, Panda AC, Munk R, Grammatikakis I, Dudekula DB, De S. Identification of HuR target circular RNAs uncovers suppression of PABPN1 translation by CircPABPN1. RNA Biol 2017; 14(3):361-9. doi: 10.1080/15476286.2017.1279788 [Crossref] [ Google Scholar]
- Liu B, Tian Y, Chen M, Shen H, Xia J, Nan J. CircUBAP2 promotes MMP9-mediated oncogenic effect via sponging miR-194-3p in hepatocellular carcinoma. Front Cell Dev Biol 2021; 9:675043. doi: 10.3389/fcell.2021.675043 [Crossref] [ Google Scholar]
- Kundu I, Varshney S, Karnati S, Naidu S. The multifaceted roles of circular RNAs in cancer hallmarks: from mechanisms to clinical implications. Mol Ther Nucleic Acids 2024; 35(3):102286. doi: 10.1016/j.omtn.2024.102286 [Crossref] [ Google Scholar]
- Li Z, Huang C, Bao C, Chen L, Lin M, Wang X. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol 2015; 22(3):256-64. doi: 10.1038/nsmb.2959 [Crossref] [ Google Scholar]
- Hu Q, Zhou T. EIciRNA-mediated gene expression: tunability and bimodality. FEBS Lett 2018; 592(20):3460-71. doi: 10.1002/1873-3468.13253 [Crossref] [ Google Scholar]
- Bose R, Ain R. Regulation of transcription by circular RNAs. Adv Exp Med Biol 2018; 1087:81-94. doi: 10.1007/978-981-13-1426-1_7 [Crossref] [ Google Scholar]
- Lu D, Xu AD. Mini review: circular RNAs as potential clinical biomarkers for disorders in the central nervous system. Front Genet 2016; 7:53. doi: 10.3389/fgene.2016.00053 [Crossref] [ Google Scholar]
- Zlotorynski E. Splicing: Going in circles. Nat Rev Genet 2018; 19(2):64-5. doi: 10.1038/nrg.2017.111 [Crossref] [ Google Scholar]
- Yang Z, He T, Chen Q. The Roles of CircRNAs in Regulating Muscle Development of Livestock Animals. Front Cell Dev Biol 2021; 9:619329. doi: 10.3389/fcell.2021.619329 [Crossref] [ Google Scholar]
- Zhang J, Chen Z, Liu X, Yang C, Xie D. Gain of circBRAF Represses Glioma Progression by Regulating miR-1290/FBXW7 Axis. Neurochemical research 2021; 46:(5), 1203-1213. doi: 10.1007/s11064-021-03259-4 [Crossref] [ Google Scholar]
- Shao T, Pan YH, Xiong XD. Circular RNA: an important player with multiple facets to regulate its parental gene expression. Mol Ther Nucleic Acids 2021; 23:369-76. doi: 10.1016/j.omtn.2020.11.008 [Crossref] [ Google Scholar]
- Granados-Riveron JT, Aquino-Jarquin G. The complexity of the translation ability of circRNAs. BiochimBiophys Acta 2016; 1859(10):1245-51. doi: 10.1016/j.bbagrm.2016.07.009 [Crossref] [ Google Scholar]
- Legnini I, Di Timoteo G, Rossi F, Morlando M, Briganti F, Sthandier O, et al. Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis. Mol Cell 2017;66(1):22-37.e9. doi: 10.1016/j.molcel.2017.02.017.
- Zhang C, Ma L, Niu Y, Wang Z, Xu X, Li Y. Circular RNA in lung cancer research: biogenesis, functions, and roles. Int J Biol Sci 2020; 16(5):803-14. doi: 10.7150/ijbs.39212 [Crossref] [ Google Scholar]
- Koch L. RNA: translated circular RNAs. Nat Rev Genet 2017; 18(5):272-3. doi: 10.1038/nrg.2017.27 [Crossref] [ Google Scholar]
- Li I, Chen YG. Emerging roles of circular RNAs in innate immunity. CurrOpin Immunol 2021; 68:107-15. doi: 10.1016/j.coi.2020.10.010 [Crossref] [ Google Scholar]
- Yu T, Wang Y, Fan Y, Fang N, Wang T, Xu T. circRNAs in cancer metabolism: a review. J Hematol Oncol 2019; 12(1):90. doi: 10.1186/s13045-019-0776-8 [Crossref] [ Google Scholar]
- Bhat AA, Gupta G, Dahiya R, Thapa R, Gahtori A, Shahwan M. circRNAs: pivotal modulators of TGF-β signalling in cancer pathogenesis. Noncoding RNA Res 2024; 9(2):277-87. doi: 10.1016/j.ncrna.2024.01.013 [Crossref] [ Google Scholar]
- Pandey PR, Munk R, Kundu G, De S, Abdelmohsen K, Gorospe M. Methods for analysis of circular RNAs. Wiley Interdiscip Rev RNA 2020; 11(1):e1566. doi: 10.1002/wrna.1566 [Crossref] [ Google Scholar]
- Gao Y, Zhao F. Computational strategies for exploring circular RNAs. Trends Genet 2018; 34(5):389-400. doi: 10.1016/j.tig.2017.12.016 [Crossref] [ Google Scholar]
- Zhang J, Hou L, Zuo Z, Ji P, Zhang X, Xue Y. Comprehensive profiling of circular RNAs with nanopore sequencing and CIRI-long. Nat Biotechnol 2021; 39(7):836-45. doi: 10.1038/s41587-021-00842-6 [Crossref] [ Google Scholar]
- Li S, Teng S, Xu J, Su G, Zhang Y, Zhao J. Microarray is an efficient tool for circRNA profiling. Brief Bioinform 2019; 20(4):1420-33. doi: 10.1093/bib/bby006 [Crossref] [ Google Scholar]
- Szabo L, Salzman J. Detecting circular RNAs: bioinformatic and experimental challenges. Nat Rev Genet 2016; 17(11):679-92. doi: 10.1038/nrg.2016.114 [Crossref] [ Google Scholar]
- Schneider T, Schreiner S, Preußer C, Bindereif A, Rossbach O. Northern blot analysis of circular RNAs. Methods Mol Biol 2018; 1724:119-33. doi: 10.1007/978-1-4939-7562-4_10 [Crossref] [ Google Scholar]
- Panda AC, Gorospe M. Detection and analysis of circular RNAs by RT-PCR. Bio Protoc 2018; 8(6):e2775. doi: 10.21769/BioProtoc.2775 [Crossref] [ Google Scholar]
- Li T, Shao Y, Fu L, Xie Y, Zhu L, Sun W. Plasma circular RNA profiling of patients with gastric cancer and their droplet digital RT-PCR detection. J Mol Med (Berl) 2018; 96(1):85-96. doi: 10.1007/s00109-017-1600-y [Crossref] [ Google Scholar]
- Chen DF, Zhang LJ, Tan K, Jing Q. Application of droplet digital PCR in quantitative detection of the cell-free circulating circRNAs. BiotechnolBiotechnol Equip 2018; 32(1):116-23. doi: 10.1080/13102818.2017.1398596 [Crossref] [ Google Scholar]
- Zirkel A, Papantonis A. Detecting circular RNAs by RNA fluorescence in situ hybridization. Methods Mol Biol 2018; 1724:69-75. doi: 10.1007/978-1-4939-7562-4_6 [Crossref] [ Google Scholar]
- Dudekula DB, Panda AC, Grammatikakis I, De S, Abdelmohsen K, Gorospe M. CircInteractome: a web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol 2016; 13(1):34-42. doi: 10.1080/15476286.2015.1128065 [Crossref] [ Google Scholar]
- Suzuki H, Tsukahara T. A view of pre-mRNA splicing from RNase R resistant RNAs. Int J Mol Sci 2014; 15(6):9331-42. doi: 10.3390/ijms15069331 [Crossref] [ Google Scholar]
- Xie R, Zhang Y, Zhang J, Li J, Zhou X. The role of circular RNAs in immune-related diseases. Front Immunol 2020; 11:545. doi: 10.3389/fimmu.2020.00545 [Crossref] [ Google Scholar]
- Xiao Y, Qiu M, Tan C, Huang W, Hu S, Jiang X. Systematic analysis of circRNA biomarkers for diagnosis, prognosis and therapy in colorectal cancer. Front Genet 2022; 13:938672. doi: 10.3389/fgene.2022.938672 [Crossref] [ Google Scholar]
- Gao L, Fan J, He J, Fan W, Che X, Wang X. Circular RNA as diagnostic and prognostic biomarkers in hematological malignancies: systematic review. Technol Cancer Res Treat 2024; 23:15330338241285149. doi: 10.1177/15330338241285149 [Crossref] [ Google Scholar]
- Wang M, Xie F, Lin J, Zhao Y, Zhang Q, Liao Z. Diagnostic and prognostic value of circulating circRNAs in cancer. Front Med (Lausanne) 2021; 8:649383. doi: 10.3389/fmed.2021.649383 [Crossref] [ Google Scholar]
- Yang Q, Li F, He AT, Yang BB. Circular RNAs: expression, localization, and therapeutic potentials. Mol Ther 2021; 29(5):1683-702. doi: 10.1016/j.ymthe.2021.01.018 [Crossref] [ Google Scholar]
- Xu C, Jun E, Okugawa Y, Toiyama Y, Borazanci E, Bolton J, et al. A circulating panel of circRNA biomarkers for the noninvasive and early detection of pancreatic ductal adenocarcinoma. Gastroenterology 2024;166(1):178-90.e16. doi: 10.1053/j.gastro.2023.09.050.
- Qin M, Liu G, Huo X, Tao X, Sun X, Ge Z. Hsa_circ_0001649: a circular RNA and potential novel biomarker for hepatocellular carcinoma. Cancer Biomark 2016; 16(1):161-9. doi: 10.3233/cbm-150552 [Crossref] [ Google Scholar]
- Zhang ZC, Guo XL, Li X. The novel roles of circular RNAs in metabolic organs. Genes Dis 2018; 5(1):16-23. doi: 10.1016/j.gendis.2017.12.002 [Crossref] [ Google Scholar]
- Pan Y, Mao Y, Jin R, Jiang L. Crosstalk between the Notch signaling pathway and non-coding RNAs in gastrointestinal cancers. Oncol Lett 2018; 15(1):31-40. doi: 10.3892/ol.2017.7294 [Crossref] [ Google Scholar]
- Chen X, Chen RX, Wei WS, Li YH, Feng ZH, Tan L. PRMT5 circular RNA promotes metastasis of urothelial carcinoma of the bladder through sponging miR-30c to induce epithelial-mesenchymal transition. Clin Cancer Res 2018; 24(24):6319-30. doi: 10.1158/1078-0432.Ccr-18-1270 [Crossref] [ Google Scholar]
- Yu CY, Kuo HC. The emerging roles and functions of circular RNAs and their generation. J Biomed Sci 2019; 26(1):29. doi: 10.1186/s12929-019-0523-z [Crossref] [ Google Scholar]
- Zhao SY, Wang J, Ouyang SB, Huang ZK, Liao L. Salivary circular RNAs hsa_circ_0001874 and hsa_circ_0001971 as novel biomarkers for the diagnosis of oral squamous cell carcinoma. Cell PhysiolBiochem 2018; 47(6):2511-21. doi: 10.1159/000491624 [Crossref] [ Google Scholar]
- Yin WB, Yan MG, Fang X, Guo JJ, Xiong W, Zhang RP. Circulating circular RNA hsa_circ_0001785 acts as a diagnostic biomarker for breast cancer detection. Clin Chim Acta 2018; 487:363-8. doi: 10.1016/j.cca.2017.10.011 [Crossref] [ Google Scholar]
- Gong Y, Mao J, Wu D, Wang X, Li L, Zhu L. Circ-ZEB133 promotes the proliferation of human HCC by sponging miR-200a-3p and upregulating CDK6. Cancer Cell Int 2018; 18:116. doi: 10.1186/s12935-018-0602-3 [Crossref] [ Google Scholar]
- Shang X, Li G, Liu H, Li T, Liu J, Zhao Q. Comprehensive circular RNA profiling reveals that hsa_circ_0005075, a new circular RNA biomarker, is involved in hepatocellular crcinoma development. Medicine (Baltimore) 2016; 95(22):e3811. doi: 10.1097/md.0000000000003811 [Crossref] [ Google Scholar]
- Xuan L, Qu L, Zhou H, Wang P, Yu H, Wu T. Circular RNA: a novel biomarker for progressive laryngeal cancer. Am J Transl Res 2016; 8(2):932-9. [ Google Scholar]
- Xu ZQ, Yang MG, Liu HJ, Su CQ. Circular RNA hsa_circ_0003221 (circPTK2) promotes the proliferation and migration of bladder cancer cells. J Cell Biochem 2018; 119(4):3317-25. doi: 10.1002/jcb.26492 [Crossref] [ Google Scholar]
- Wang X, Zhang Y, Huang L, Zhang J, Pan F, Li B. Decreased expression of hsa_circ_001988 in colorectal cancer and its clinical significances. Int J Clin Exp Pathol 2015; 8(12):16020-5. [ Google Scholar]
- Li P, Chen S, Chen H, Mo X, Li T, Shao Y. Using circular RNA as a novel type of biomarker in the screening of gastric cancer. Clin Chim Acta 2015; 444:132-6. doi: 10.1016/j.cca.2015.02.018 [Crossref] [ Google Scholar]
- Dong Y, Gao Q, Chen Y, Zhang Z, Du Y, Liu Y. Identification of circRNA signature associated with tumor immune infiltration to predict therapeutic efficacy of immunotherapy. Nat Commun 2023; 14(1):2540. doi: 10.1038/s41467-023-38232-y [Crossref] [ Google Scholar]
- Li LX, Hao Y, Dong L, Qiao ZQ, Yang SC, Chen YD. Circular RNAs as biomarkers in breast cancer diagnosis, prognosis, molecular types, metastasis and drug resistance. Technol Cancer Res Treat 2025; 24:15330338251328500. doi: 10.1177/15330338251328500 [Crossref] [ Google Scholar]
- Wu X, Shi M, Lian Y, Zhang H. Exosomal circRNAs as promising liquid biopsy biomarkers for glioma. Front Immunol 2023; 14:1039084. doi: 10.3389/fimmu.2023.1039084 [Crossref] [ Google Scholar]
- Yi J, Wang L, Hu GS, Zhang YY, Du J, Ding JC. CircPVT1 promotes ER-positive breast tumorigenesis and drug resistance by targeting ESR1 and MAVS. EMBO J 2023; 42(10):e112408. doi: 10.15252/embj.2022112408 [Crossref] [ Google Scholar]
- Tierno D, Grassi G, Zanconati F, Dapas B, Scaggiante B. Plasma circular RNAs as biomarkers for breast cancer. Biomedicines 2024; 12(4):875. doi: 10.3390/biomedicines12040875 [Crossref] [ Google Scholar]
- Sarkar D, Diermeier SD. Circular RNAs: potential applications as therapeutic targets and biomarkers in breast cancer. Noncoding RNA 2021; 7(1):2. doi: 10.3390/ncrna7010002 [Crossref] [ Google Scholar]
- Yan Y, Su M, Qin B. CircHIPK3 promotes colorectal cancer cells proliferation and metastasis via modulating of miR-1207-5p/FMNL2 signal. BiochemBiophys Res Commun 2020; 524(4):839-46. doi: 10.1016/j.bbrc.2020.01.055 [Crossref] [ Google Scholar]
- Li Y, Zheng F, Xiao X, Xie F, Tao D, Huang C. CircHIPK3 sponges miR-558 to suppress heparanase expression in bladder cancer cells. EMBO Rep 2017; 18(9):1646-59. doi: 10.15252/embr.201643581 [Crossref] [ Google Scholar]
- Li F, Zhang L, Li W, Deng J, Zheng J, An M. Circular RNA ITCH has inhibitory effect on ESCC by suppressing the Wnt/β-catenin pathway. Oncotarget 2015; 6(8):6001-13. doi: 10.18632/oncotarget.3469 [Crossref] [ Google Scholar]
- Su Y, Lv X, Yin W, Zhou L, Hu Y, Zhou A. circRNA Cdr1as functions as a competitive endogenous RNA to promote hepatocellular carcinoma progression. Aging (Albany NY) 2019; 11(19):8183-203. doi: 10.18632/aging.102312 [Crossref] [ Google Scholar]
- Sang M, Meng L, Liu S, Ding P, Chang S, Ju Y. Circular RNA ciRS-7 maintains metastatic phenotypes as a ceRNA of miR-1299 to target MMPs. Mol Cancer Res 2018; 16(11):1665-75. doi: 10.1158/1541-7786.Mcr-18-0284 [Crossref] [ Google Scholar]
- Xie X, Sun FK, Huang X, Wang CH, Dai J, Zhao JP. A circular RNA, circSMARCA5, inhibits prostate cancer proliferative, migrative, and invasive capabilities via the miR-181b-5p/miR-17-3p-TIMP3 axis. Aging (Albany NY) 2021; 13(15):19908-19. doi: 10.18632/aging.203408 [Crossref] [ Google Scholar]
- Yang Y, Gao X, Zhang M, Yan S, Sun C, Xiao F. Novel role of FBXW7 circular RNA in repressing glioma tumorigenesis. J Natl Cancer Inst 2018; 110(3):304-15. doi: 10.1093/jnci/djx166 [Crossref] [ Google Scholar]
- Zhang Y, Zhao H, Zhang L. Identification of the tumor-suppressive function of circular RNA FOXO3 in non-small cell lung cancer through sponging miR-155. Mol Med Rep 2018; 17(6):7692-700. doi: 10.3892/mmr.2018.8830 [Crossref] [ Google Scholar]
- Kong Z, Wan X, Lu Y, Zhang Y, Huang Y, Xu Y. Circular RNA circFOXO3 promotes prostate cancer progression through sponging miR-29a-3p. J Cell Mol Med 2020; 24(1):799-813. doi: 10.1111/jcmm.14791 [Crossref] [ Google Scholar]
- Fang G, Ye BL, Hu BR, Ruan XJ, Shi YX. circRNA_100290 promotes colorectal cancer progression through miR-516b-induced downregulation of FZD4 expression and Wnt/β-catenin signaling. BiochemBiophys Res Commun 2018; 504(1):184-9. doi: 10.1016/j.bbrc.2018.08.152 [Crossref] [ Google Scholar]
- Li H, Xue S, Zhang X, Li F, Bei S, Feng L. circRNA PVT1 modulated cell migration and invasion through Epithelial-Mesenchymal Transition (EMT) mediation in gastric cancer through miR-423-5p/Smad3 pathway. Regen Ther 2022; 21:25-33. doi: 10.1016/j.reth.2022.02.003 [Crossref] [ Google Scholar]
- Liu YP, Wan J, Long F, Tian J, Zhang C. circPVT1 facilitates invasion and metastasis by regulating miR-205-5p/c-FLIP axis in osteosarcoma. Cancer Manag Res 2020; 12:1229-40. doi: 10.2147/cmar.S231872 [Crossref] [ Google Scholar]
- Sang Y, Chen B, Song X, Li Y, Liang Y, Han D. circRNA_0025202 regulates tamoxifen sensitivity and tumor progression via regulating the miR-182-5p/FOXO3a axis in breast cancer. Mol Ther 2019; 27(9):1638-52. doi: 10.1016/j.ymthe.2019.05.011 [Crossref] [ Google Scholar]
- He R, Liu P, Xie X, Zhou Y, Liao Q, Xiong W. circGFRA1 and GFRA1 act as ceRNAs in triple negative breast cancer by regulating miR-34a. J Exp Clin Cancer Res 2017; 36(1):145. doi: 10.1186/s13046-017-0614-1 [Crossref] [ Google Scholar]
- Zhang PF, Gao C, Huang XY, Lu JC, Guo XJ, Shi GM. Cancer cell-derived exosomal circUHRF1 induces natural killer cell exhaustion and may cause resistance to anti-PD1 therapy in hepatocellular carcinoma. Mol Cancer 2020; 19(1):110. doi: 10.1186/s12943-020-01222-5 [Crossref] [ Google Scholar]
- Qiu M, Xia W, Chen R, Wang S, Xu Y, Ma Z. The circular RNA circPRKCI promotes tumor growth in lung adenocarcinoma. Cancer Res 2018; 78(11):2839-51. doi: 10.1158/0008-5472.Can-17-2808 [Crossref] [ Google Scholar]
- Wang S, Xue X, Wang R, Li X, Li Q, Wang Y. CircZNF609 promotes breast cancer cell growth, migration, and invasion by elevating p70S6K1 via sponging miR-145-5p. Cancer Manag Res 2018; 10:3881-90. doi: 10.2147/cmar.S174778 [Crossref] [ Google Scholar]
- Rinaldi C, Wood MJA. Antisense oligonucleotides: the next frontier for treatment of neurological disorders. Nat Rev Neurol 2018; 14(1):9-21. doi: 10.1038/nrneurol.2017.148 [Crossref] [ Google Scholar]
- Cao SQ, Xue ST, Li WJ, Hu GS, Wu ZG, Zheng JC. CircHIPK3 regulates fatty acid metabolism through miR-637/FASN axis to promote esophageal squamous cell carcinoma. Cell Death Discov 2024; 10(1):110. doi: 10.1038/s41420-024-01881-z [Crossref] [ Google Scholar]
- Pollak AJ, Zhao L, Crooke ST. Systematic analysis of chemical modifications of phosphorothioate antisense oligonucleotides that modulate their innate immune response. Nucleic Acid Ther 2023; 33(2):95-107. doi: 10.1089/nat.2022.0067 [Crossref] [ Google Scholar]
- Katti A, Diaz BJ, Caragine CM, Sanjana NE, Dow LE. CRISPR in cancer biology and therapy. Nat Rev Cancer 2022; 22(5):259-79. doi: 10.1038/s41568-022-00441-w [Crossref] [ Google Scholar]
- Zhang J, Luo Z, Zheng Y, Duan M, Qiu Z, Huang C. circRNA as an Achilles heel of cancer: characterization, biomarker and therapeutic modalities. J Transl Med 2024; 22(1):752. doi: 10.1186/s12967-024-05562-4 [Crossref] [ Google Scholar]
- Wen L, Zhao C, Song J, Ma L, Ruan J, Xia X. CRISPR/Cas9-mediated TERT disruption in cancer cells. Int J Mol Sci 2020; 21(2):653. doi: 10.3390/ijms21020653 [Crossref] [ Google Scholar]
- Zhang Y, Nguyen TM, Zhang XO, Wang L, Phan T, Clohessy JG. Optimized RNA-targeting CRISPR/Cas13d technology outperforms shRNA in identifying functional circRNAs. Genome Biol 2021; 22(1):41. doi: 10.1186/s13059-021-02263-9 [Crossref] [ Google Scholar]
- Zhang M, Guo R, Yuan Z, Wang H. Lipid nanoparticle (LNP)-a vector suitable for evolving therapies for advanced hepatocellular carcinoma (HCC). Glob Chall 2025; 9(1):2400217. doi: 10.1002/gch2.202400217 [Crossref] [ Google Scholar]
- Yao Y, Zhou Y, Liu L, Xu Y, Chen Q, Wang Y. Nanoparticle-based drug delivery in cancer therapy and its role in overcoming drug resistance. Front Mol Biosci 2020; 7:193. doi: 10.3389/fmolb.2020.00193 [Crossref] [ Google Scholar]
- Cai J, Liu Z, Chen S, Zhang J, Li H, Wang X. Engineered circular RNA-based DLL3-targeted CAR-T therapy for small cell lung cancer. Exp Hematol Oncol 2025; 14(1):35. doi: 10.1186/s40164-025-00625-8 [Crossref] [ Google Scholar]
- Ning H, Jiang Y, Li B, Ren J, Ran A, Li W. Targeted delivery of circPDHK1 siRNA via aptamer functionalized lipid nanoparticles inhibits ccRCC growth and migration. Int J Pharm 2025; 677:125666. doi: 10.1016/j.ijpharm.2025.125666 [Crossref] [ Google Scholar]
- You S, Luo Z, Cheng N, Wu M, Lai Y, Wang F. Magnetically responsive nanoplatform targeting circRNA circ_0058051 inhibits hepatocellular carcinoma progression. Drug DelivTransl Res 2023; 13(3):782-94. doi: 10.1007/s13346-022-01237-z [Crossref] [ Google Scholar]
- Shu G, Lu X, Pan Y, Cen J, Huang K, Zhou M. Exosomal circSPIRE1 mediates glycosylation of E-cadherin to suppress metastasis of renal cell carcinoma. Oncogene 2023; 42(22):1802-20. doi: 10.1038/s41388-023-02678-7 [Crossref] [ Google Scholar]
- Gopikrishnan M, R HC, R G, Ashour HM, Pintus G, Hammad M. Therapeutic and diagnostic applications of exosomal circRNAs in breast cancer. FunctIntegr Genomics 2023; 23(2):184. doi: 10.1007/s10142-023-01083-3 [Crossref] [ Google Scholar]
- Motoyama K, Inoue H, Mimori K, Tanaka F, Kojima K, Uetake H. Clinicopathological and prognostic significance of PDCD4 and microRNA-21 in human gastric cancer. Int J Oncol 2010; 36(5):1089-95. doi: 10.3892/ijo_00000590 [Crossref] [ Google Scholar]
- Müller S, Wedler A, Breuer J, Glaß M, Bley N, Lederer M. Synthetic circular miR-21 RNA decoys enhance tumor suppressor expression and impair tumor growth in mice. NAR Cancer 2020; 2(3):zcaa014. doi: 10.1093/narcan/zcaa014 [Crossref] [ Google Scholar]
- Yang C, Yuan W, Yang X, Li P, Wang J, Han J. Circular RNA circ-ITCH inhibits bladder cancer progression by sponging miR-17/miR-224 and regulating p21, PTEN expression. Mol Cancer 2018; 17(1):19. doi: 10.1186/s12943-018-0771-7 [Crossref] [ Google Scholar]
- Ameli-Mojarad M, Ameli-Mojarad M, Hadizadeh M, Young C, Babini H, Nazemalhosseini-Mojarad E. The effective function of circular RNA in colorectal cancer. Cancer Cell Int 2021; 21(1):496. doi: 10.1186/s12935-021-02196-0 [Crossref] [ Google Scholar]
- Huang G, Zhu H, Shi Y, Wu W, Cai H, Chen X. cir-ITCH plays an inhibitory role in colorectal cancer by regulating the Wnt/β-catenin pathway. PLoS One 2015; 10(6):e0131225. doi: 10.1371/journal.pone.0131225 [Crossref] [ Google Scholar]
- Peng Y, Wang HH. cir-ITCH inhibits gastric cancer migration, invasion and proliferation by regulating the Wnt/β-catenin pathway. Sci Rep 2020; 10(1):17443. doi: 10.1038/s41598-020-74452-8 [Crossref] [ Google Scholar]
- Yuan Z, Huang S, Jin X, Li S. Circular RNAs in cardiovascular diseases: molecular mechanisms, therapeutic advances, and innovations. Genes (Basel) 2024; 15(11):1423. doi: 10.3390/genes15111423 [Crossref] [ Google Scholar]
- Barbagallo D, Caponnetto A, Cirnigliaro M, Brex D, Barbagallo C, D’Angeli F. CircSMARCA5 inhibits migration of glioblastoma multiforme cells by regulating a molecular axis involving splicing factors SRSF1/SRSF3/PTB. Int J Mol Sci 2018; 19(2):480. doi: 10.3390/ijms19020480 [Crossref] [ Google Scholar]
- Du WW, Yang W, Liu E, Yang Z, Dhaliwal P, Yang BB. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res 2016; 44(6):2846-58. doi: 10.1093/nar/gkw027 [Crossref] [ Google Scholar]
- Meng L, Wu H, Wu J, Ding P, He J, Sang M. Mechanisms of immune checkpoint inhibitors: insights into the regulation of circular RNAS involved in cancer hallmarks. Cell Death Dis 2024; 15(1):3. doi: 10.1038/s41419-023-06389-5 [Crossref] [ Google Scholar]
- Hong W, Xue M, Jiang J, Zhang Y, Gao X. Circular RNA circ-CPA4/ let-7 miRNA/PD-L1 axis regulates cell growth, stemness, drug resistance and immune evasion in non-small cell lung cancer (NSCLC). J Exp Clin Cancer Res 2020; 39(1):149. doi: 10.1186/s13046-020-01648-1 [Crossref] [ Google Scholar]
- Yang K, Zhang J, Bao C. Exosomal circEIF3K from cancer-associated fibroblast promotes colorectal cancer (CRC) progression via miR-214/PD-L1 axis. BMC Cancer 2021; 21(1):933. doi: 10.1186/s12885-021-08669-9 [Crossref] [ Google Scholar]
- Li S, Li X, Xue W, Zhang L, Yang LZ, Cao SM. Screening for functional circular RNAs using the CRISPR-Cas13 system. Nat Methods 2021; 18(1):51-9. doi: 10.1038/s41592-020-01011-4 [Crossref] [ Google Scholar]
- Williford JM, Wu J, Ren Y, Archang MM, Leong KW, Mao HQ. Recent advances in nanoparticle-mediated siRNA delivery. Annu Rev Biomed Eng 2014; 16:347-70. doi: 10.1146/annurev-bioeng-071813-105119 [Crossref] [ Google Scholar]
- Sani A, Cao C, Cui D. Toxicity of gold nanoparticles (AuNPs): a review. BiochemBiophys Rep 2021; 26:100991. doi: 10.1016/j.bbrep.2021.100991 [Crossref] [ Google Scholar]
- Zwolsman R, Darwish YB, Kluza E, van der Meel R. Engineering lipid nanoparticles for mRNA immunotherapy. Wiley Interdiscip Rev NanomedNanobiotechnol 2025; 17(2):e70007. doi: 10.1002/wnan.70007 [Crossref] [ Google Scholar]
- He AT, Liu J, Li F, Yang BB. Targeting circular RNAs as a therapeutic approach: current strategies and challenges. Signal Transduct Target Ther 2021; 6(1):185. doi: 10.1038/s41392-021-00569-5 [Crossref] [ Google Scholar]
- Holdt LM, Kohlmaier A, Teupser D. Circular RNAs as therapeutic agents and targets. Front Physiol 2018; 9:1262. doi: 10.3389/fphys.2018.01262 [Crossref] [ Google Scholar]
- Zhang Y, Wang Y, Su X, Wang P, Lin W. The value of circulating circular RNA in cancer diagnosis, monitoring, prognosis, and guiding treatment. Front Oncol 2021; 11:736546. doi: 10.3389/fonc.2021.736546 [Crossref] [ Google Scholar]
- Cai J, Qiu Z, Chi-Shing Cho W, Liu Z, Chen S, Li H. Synthetic circRNA therapeutics: innovations, strategies, and future horizons. MedComm (2020) 2024; 5(11):e720. doi: 10.1002/mco2.720 [Crossref] [ Google Scholar]
- Márton É, Varga A, Domoszlai D, Buglyó G, Balázs A, Penyige A. Non-coding RNAs in cancer: structure, function, and clinical application. Cancers (Basel) 2025; 17(4):579. doi: 10.3390/cancers17040579 [Crossref] [ Google Scholar]
- Zhang X, Yuan Y, Wang X, Wang H, Zhang L, He J. CircWHSC1 (CircNSD2): a novel circular RNA in multiple cancers. Clin Med Insights Oncol 2024; 18:11795549241254781. doi: 10.1177/11795549241254781 [Crossref] [ Google Scholar]