Advanced pharmaceutical bulletin. 15(4):700-716.
doi: 10.34172/apb.025.46020
Review Article
The Role of Extracellular Vesicles in Neuropsychiatric Disorders
Behnaz Mirzaahmadi Data curation, Funding acquisition, Investigation, Methodology, Resources, 1 
Reza Rahbarghazi Formal analysis, Software, Validation, Visualization, 2
Mohammad Karimipour Conceptualization, Project administration, Supervision, 3, *
Author information:
1Student Research Committee, Tabriz University of Medical Sciences, Tabriz, Iran
2Department of Applied Cell Sciences, Faculty of Advanced Medical Sciences, Tabriz University of Medical Sciences, Tabriz, Iran
3Department of Anatomical Sciences, Tabriz University of Medical Sciences, Tabriz, Iran
Abstract
Purpose:
There are effective treatments available for neuropsychiatric disorders; however, numerous factors such as misdiagnosis, delayed diagnosis, varying types of onsets and progression of disorder, as well as long timeframes between diagnosis and treatment, all serve to limit the ability of these treatments to reduce symptoms. This article discusses the unique theragnostic aspects of extracellular vesicles (EVs) in relation to these clinical considerations.
Methods:
An extensive review of the literature was conducted to examine recent findings about EVs and their potential role in early diagnosis and targeted treatment of neuropsychiatric disorders, to examine the biological aspects, diagnostic and therapeutic capacity.
Results:
Pathological states radically change the composition of EVs and their cargo delivery, thereby providing a dynamic disease state. EVs can cross biological barriers, have systemic distribution, provide low immunogenicity and toxicity, and all serve to represent the beginnings of identifying early biomarkers and future methods for delivering therapy needed to administer neuroactive drugs/delivery methods.
Conclusion:
In summary, EVs have potential as a theragnostic platform for neuropsychiatric diseases. Because they offer passive, non-invasive, and actionable diagnostic insights, EVs can be applied to therapeutically manage neuropsychiatric disorders, which could provide a solution to issues faced in clinical practice, such as late-stage diagnosis and poor treatment compliance. To fully understand the prospect of EVs in neuropsychiatric practice, more translational research and clinical validation are required.
Keywords: Extracellular vesicles, Neuropsychiatric disorders, Therapeutic effects
Copyright and License Information
© 2025 The Author (s).
This is an Open Access article distributed under the terms of the Creative Commons Attribution (CC BY), which permits unrestricted use, distribution, and reproduction in any medium, as long as the original authors and source are cited. No permission is required from the authors or the publishers.
Funding Statement
The research protocol was approved & Supported by the Student Research Committee, Tabriz University of Medical Sciences (Grant No: 72817).
Introduction
Neuropsychiatric disorders include a range of complex and debilitating conditions that significantly impact patients’ lives with heavy socioeconomic burdens.1 Among neuropsychiatric disorders, major depressive disorder (MDD), schizophrenia (SCZ), bipolar disorder (BD), and Alzheimer’s disease (AD) are the most common in the human population. Still, their etiology remains poorly understood due to ethical issues, practical challenges, the complex biology of higher brain function, and the weak validation of animal models.2 Neuroinflammation, oxidative stress, neurodegeneration, and the modulation of specific signaling pathways and neurotransmitters are the primary causes of the onset and progression of these disorders.3,4 Currently, clinical manifestations, symptoms, and cognitive tests are still used to diagnose individuals suspected of having the condition.5 Despite recent progress, the absence of accurate objective signs or biomarkers has led to the possibility of misdiagnosis and delays in initiating treatment protocols.6 To develop effective therapeutic protocols, understanding the underlying cellular mechanisms of neuropsychiatric disorders is crucial. Consequently, ongoing studies have focused on identifying new targets for drug development within the central nervous system (CNS).
The ability of extracellular vesicles (EVs) to provide important diagnostic highlights or markers, while simultaneously delivering the therapeutic agent directly to the targeted site, is a significant benefit for the patient, allowing for the integration of both diagnostic and potential treatment delivery interventions in a targeted manner. The use of this system would also provide physicians with real-time evidence of the intervention’s effectiveness, enabling them to make adjustments based on patient-specific needs.7 This review will examine both aspects of EV function by providing a summary of key biomarkers with diagnostic potential in neuropsychiatric disorders and discussing the potential adoption of EVs as therapeutic agents in modulating disease processes (Figure 1). Importantly, we will consider the integration of these important and innovative diagnostics and therapeutic functions to be a proper theragnostic within clinical neuropsychiatry, despite conflicting evidence.
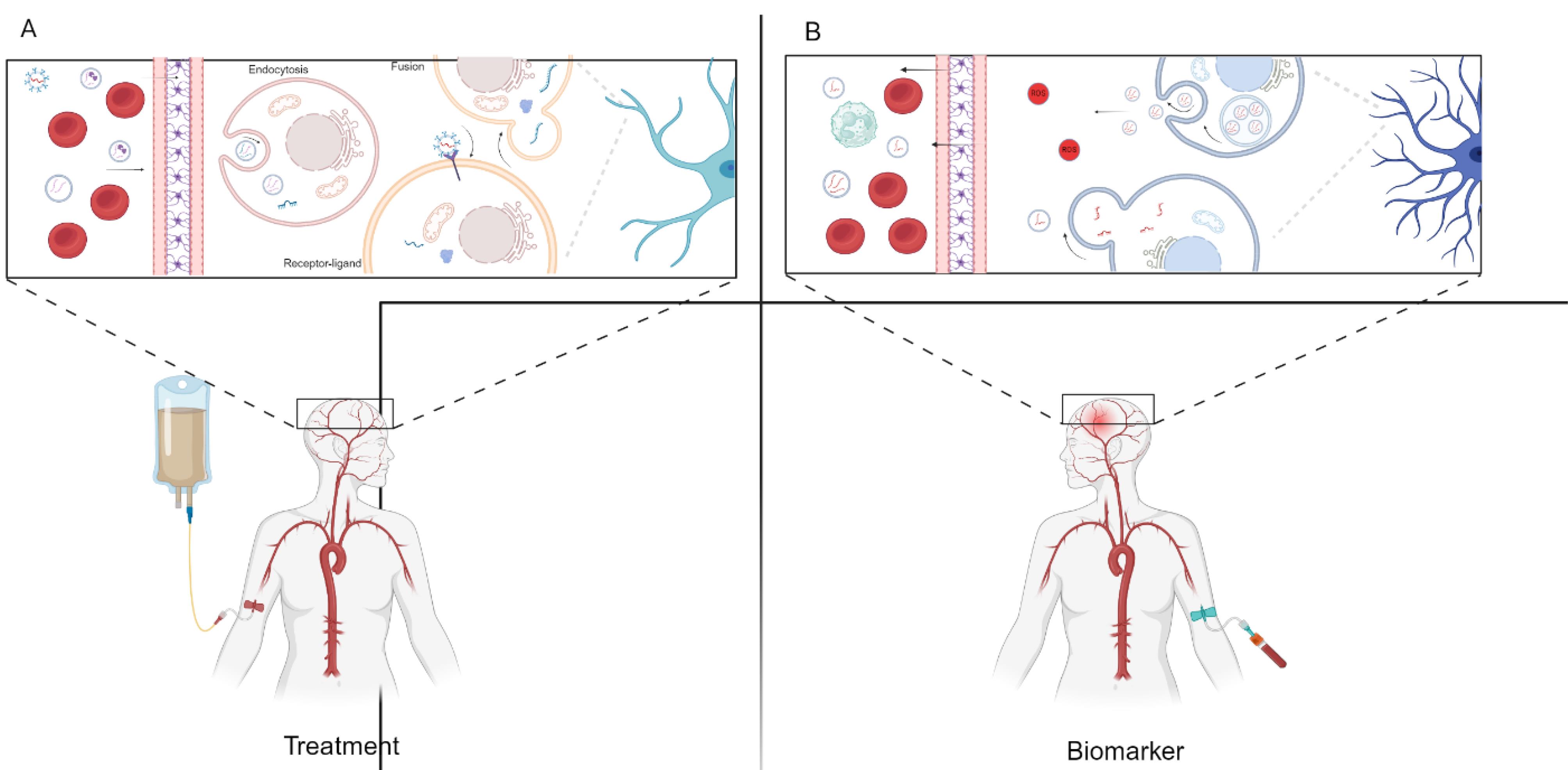
Figure 1.
EVs have essential roles as biomarkers and in treatment. A. EVs hold promise as therapeutic agents for neuropsychiatric disorders, offering targeted delivery of therapeutic cargo to affected brain regions. B. EVs offer diagnostic potential, serving as biomarkers for early detection and monitoring of neuropsychiatric disorders
.
EVs have essential roles as biomarkers and in treatment. A. EVs hold promise as therapeutic agents for neuropsychiatric disorders, offering targeted delivery of therapeutic cargo to affected brain regions. B. EVs offer diagnostic potential, serving as biomarkers for early detection and monitoring of neuropsychiatric disorders
EVs in CNS
EVs are an emerging field of biological research with the potential for treating neuropsychiatric disorders.8 EVs are endosomal, small, membrane-bound structures released by almost every cell into the extracellular matrix (ECM). EVs, especially exosomes (Exos), have garnered considerable attention due to their ability to transport signaling molecules, including proteins, lipids, and nucleic acids, between cells.9 These vesicles have been shown to play roles in neuroplasticity, neuroprotection, and improvement of neural dysfunction.10 Emerging data suggest that EVs have substantial roles in the pathophysiology and physiology of neuropsychiatric disorders.11 Within the CNS, EVs are produced by various cell types, including glial cells and neurons.12 For instance, as part of neuroimmune interactions, inflammatory astrocytes can produce EVs that can damage neurons.13 Of note, these particles can cross the biological barriers, such as the blood-brain barrier (BBB). It was suggested that the juxtaposition of EVs with endothelial cells (ECs) leads to the activation of several subcellular mechanisms, such as endocytosis, micropinocytosis, phagocytosis, and direct plasma fusion, resulting in EVs crossing the BBB.14 It should not be forgotten that their cargoes may also affect how and by which mechanisms EVs cross the BBB.11 EVs are also involved in the removal of waste byproducts in the CNS, helping the host cells eliminate aggregates of neurotoxic substances and reducing drug resistance by bypassing the P-glycoprotein drug efflux pathway.15,16 Besides, EVs play crucial roles in neurodevelopment and the homeostasis of the brain’s neuronal function and secretion.17 The uptake of EVs by target neuronal cells is dependent on lipid components in their membranes, such as sphingomyelin and phosphatidylserine.18
EV biogenesis encompasses multiple molecular pathways that orchestrate the uptake of EVs, their fusion with endosomal systems, and the release of their cargo into the cytosol (Figure 2).19 In this process, EV ligands can bind to receptors on the recipient cells, activating entry mechanisms such as endocytosis, phagocytosis, and micropinocytosis.20 Following arrival in the cytosol, EV genomics, such as RNA, can alter host cell functions by changing gene expression or affecting certain signaling pathways.21 Among various genomic cargoes, EV miRNAs are valid biomarkers for detecting neuropsychiatric disorders.22 Interestingly, nearly 70% of body miRNAs are produced in the brain.23 These features demonstrate the theragnostic importance of miRNAs in the pathophysiology of several CNS pathological conditions. Other genetic elements, lncRNAs and circRNAs, also exist inside the EVs.24 LncRNAs with an average size of more than 200 nucleotides can regulate transcription, metabolism, signaling pathways, protein function, and RNA stability.25 circRNAs, covalently closed RNAs, function as protein scaffolds or miRNA sponges to regulate several cell functions.26 Among the various RNA types, circular RNAs (circRNAs) are more resistant to enzymatic degradation and thus possess longer lifespans, making them more reliable biomarkers for chronic pathologies and therapeutic targets.27
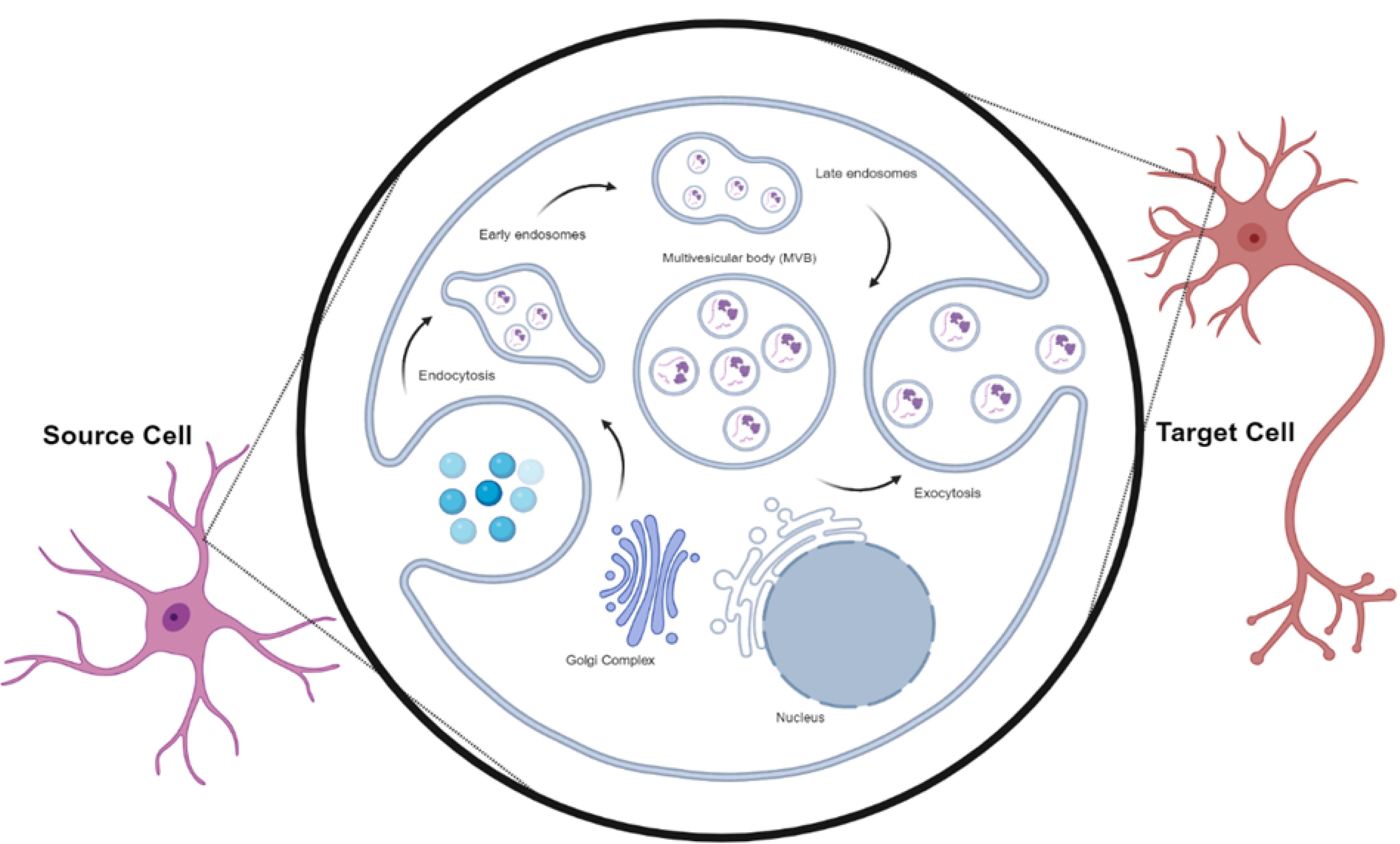
Figure 2.
Exosome biogenesis. The internalized Exos are directed into early endosomes induced through cell membrane invagination. In the following steps, the invagination of early, late endosomes, and multivesicular bodies (MVBs) leads to the generation of several intraluminal vesicles (ILVs). The fusion of MVBs with the cell membrane contributes to the release of ILVs into the ECM, which are subsequently referred to as Exos, belonging to EVs
.
Exosome biogenesis. The internalized Exos are directed into early endosomes induced through cell membrane invagination. In the following steps, the invagination of early, late endosomes, and multivesicular bodies (MVBs) leads to the generation of several intraluminal vesicles (ILVs). The fusion of MVBs with the cell membrane contributes to the release of ILVs into the ECM, which are subsequently referred to as Exos, belonging to EVs
EV biomarkers provide essential data from the parent cells. For instance, the existence of glutamate-aspartate transporters indicates that these EVs originate from glial cells.28 In the CNS, L1CAM is used to discriminate neuron EVs from other cell sources.29 In addition to their diagnostic potential, EVs can be modified and bioengineered to transfer signaling biomolecules and therapeutics, thereby influencing gene expression, translation, and signaling pathways for the treatment of specific pathological conditions.30 Specific physicochemical properties, such as a slight negative zeta potential, flexible cytoskeleton, and low immunogenicity, support their application as nanocarriers for drug delivery to the CNS.31 These properties indicate their superiority compared to synthetic nanocarriers that are cytotoxic or cleared by the reticuloendothelial system.32
For sampling, EVs can be obtained from different biofluids for various scientific purposes. Using liquid biopsy, EVs have been obtained from the postmortem brain, cerebrospinal fluid (CSF), and blood. To enrich these particles, techniques such as size exclusion chromatography, immunoisolation, differential ultracentrifugation (UC) steps, microfiltration, and microfluidic systems are available.33 However, the isolation methods can, in part but not completely, alter the structural integrity and molecular contents, and thus, there is an urgent demand to develop standardized extraction protocols to minimize unintended in vitro changes. The process used to isolate the EVs will also impact, to some degree, the sensitivity and specificity of the assay.34 UC is the standardized method, which is great in its specificity and lower in contamination, but also yields less.35 Polyethylene glycol (PEG) is a precipitation-based technique that is quicker and yields more proteins/particles, but typically co-isolates more proteins/particles, thereby lowering specificity.36 Size exclusion chromatography (SEC) balances yield with low contamination and is less damaging to vesicle structure, making this method more ideal for studies that attempt to detect sensitive biomarkers.37 Antibody-coated magnetic beads, combined with immunoaffinity-based methods, can increase the specificity and sensitivity of EV subtype separation by explicitly utilizing the surface markers of EVs; however, this approach often requires some prior knowledge of EV surface markers.38 Studies that use magnetic beads are limited by the volume of the samples, which can potentially contaminate the enriched EVs compared to other methods.37 Preanalytical variables, including biofluid (e.g., plasma, serum, cerebrospinal fluid), storage temperature, freeze-thaw cycles, and handling, will impact the integrity of EVs, thereby affecting the reliability of biomarkers and often leading to inconsistencies in results. In the examples, improper sample storage can alter or aggregate EVs, thus limiting the potential for meaningful detection and any molecular profiling or experimental use of EVs thereafter.39 If nothing else, standardizing the steps involved in EV isolation protocols and preanalytical factors can only make biomarker studies more informative.
Role of EVs in SCZ
SCZ, a severe neuropsychiatric disorder with a prevalence of about 1% of the human population, exhibits a high recurrence and disability rate.40 A major predictor of disability and functioning in SCZ is cognitive deficits.41 Globally, the life expectancy in SCZ patients ranges between 10 and 20 years, and for some patients leads to suicide.42 It has been thought that a close interaction between genetic elements and the environment underpins these conditions.4 SCZ patients are still diagnosed between adolescence and early adulthood by clinical interviews and observations, with a risk of delay and misdiagnosis when starting treatment.40 Timely diagnosis is an essential step toward recovery. Medications using antipsychotics mainly alleviate symptoms such as hallucinations, but don’t have a remarkable impact on cognitive symptoms such as learning and attention.4 Molecular investigations have revealed that inflammatory response, neurotrophic deficit, and epigenetic regulation have been implicated as potential biomarkers of SCZ.43 Several candidate molecules have been proposed as SCZ biomarkers using “omics” methods, including proteomics, transcriptomics, and metabolomics.44 Despite the existence of several proposed biomarkers for SCZ, the diagnosis of SCZ based on candidate biomarkers remains elusive. Therefore, it is crucial to understand the etiology of this disease in order to develop novel biomarkers that aid in diagnosis and prognosis. Emerging data have indicated the presence of valid signaling molecules, such as miRNAs, circRNAs, metabolites, and proteins, inside the EVs under SCZ conditions.45,46 The differential expression of 44 circRNAs involved in stress response and pathogenesis has been verified in the plasma of SCZ patients and healthy counterparts. It is believed that EV miRNAs act as epigenetic regulators during the onset of SCZ.47
SCZ patients had significantly higher levels of 11 miRNAs compared to healthy controls, as determined by genome-wide interactive analysis of blood exosome miRNA data. Among these miRNAs, hsa-miR-206 exhibited the highest levels, and animal models demonstrated that hsa-miR-206 has detrimental effects on cognitive functions by disrupting brain-derived neurotrophic factor (BDNF) mRNA and protein expression. As a result, the abnormalities in the brains of SCZ patients are associated with disturbances of neurotrophic factors.48 There is evidence that SCZ has a linkage with excessive dopaminergic activity in the mesolimbic tract, gamma-aminobutyric acid (GABA)-ergic inhibitory activities, decreased dopamine in the prefrontal cortex, and reduction of mesocortical dopaminergic neurons.49 In a study conducted by Funahashi et al, microarray analysis revealed 18 down-regulated and 13 up-regulated miRNAs in SCZ conditions linked to brain development. Of note, the non-treatment-resistant SCZ group exhibits a significant reduction in miR-675-3p expression.50 But this study has limitations that included a small sample size for the microarray cohort (there were only 9 patients and 9 controls), and the SH-SY5Y cell line used for clozapine stimulation may not entirely recapitulate the complex environment in vivo within the brain, including cell-specific effects.
The transport of misfolded proteins and neuroinflammatory cytokines by EVs from different neuronal cells can increase the possibility of SCZ. Under such conditions, EV metabolites related to glycerophospholipids, phenylalanine, tyrosine, or tryptophan have been altered.51 Goetzl et al enriched astrocyte and neuron EVs using immunoprecipitation methods from the plasma of first-episode psychotics and controls. It was found that the levels of subunits 1 and 6 of complex I and subunit 10 of complex III were remarkably decreased, and ROS levels increased in the EVs of SCZ patients. The elevation of glial fibrillary acidic protein (GFAP) in EVs is closely associated with the polarization of astrocytes to the A1 type in SCZ patients. But this study has several limitations, including the challenge of precisely isolating and identifying EVs derived purely from neural sources in peripheral blood, considering the blood EV pool is a mixture of EVs derived from many tissues and cells that hampers capacity to identify changes that are uniquely related to brain pathology, and comparatively small sample sizes or heterogeneous patient populations may restrict statistical power and generalizability.52
In another study, a noticeable difference was achieved in the plasma-derived EV lncRNAs, such as myocardial infarction-associated transcript (MIAT) and plasmacytoma variant translocation 1 (PVT1), between patients and controls.53 At varying degrees of activity, some types of miRNAs are essential in the pathophysiology of neuropsychiatric diseases. For instance, miR-132, which targets the CLOCK gene, is decreased in SCZ but increased in MDD and BD. miR-132 also has broad biological impacts, including the regulation of hippocampal neurogenesis, differentiation, axonal growth, neural plasticity, and the alleviation of memory deficits.54
Like the alteration of genetic cargoes in EVs of SCZ, the lipid profile of plasma EVs can also be modified in SCZ candidates. Along with these changes, prefrontal cortex glycerophospholipid metabolism was decreased, and blood malondialdehyde (MDA) levels were elevated. These features indicate the role of lipid peroxidation in the pathogenesis of SCZ.55 The membrane phospholipid hypothesis of SCZ describes the critical role that phospholipids play in the development of the brain and its functioning. Based on this hypothesis, abnormal levels of essential fatty acids in the peripheral blood of SCZ individuals are integral to high-risk psychosis individuals, highlighting the focus on lipid metabolism as a therapeutic target for SCZ patients.56 Due to the current limited knowledge of the role of EVs in SZC, resulting from a lack of experimental results, further research is needed to identify the pathways involved in treatment and diagnosis.
Table 1 provides a summary of several relevant biomarkers on EVs, including proteins, nucleic acids, and lipids, described in neuropsychiatric disorders.
Table 1.
Diagnostic importance of EVs in neuropsychiatric disorders
|
Ref
|
Disease
|
Species
|
Biofluid
|
EV cargo
|
Diagnostic potential
|
|
57
|
SCZ |
Human |
Plasma |
higher ADE-P-T181-tau |
Executive functioning in SCZ |
|
48
|
SCZ |
Human |
serum |
hsa-miR-206 |
Targets the BDNF mRNA |
|
48
|
SCZ |
Human |
serum |
hsa-miR145-5p, and hsa-miR-133a-3p |
2-fold increase in SCZ |
|
58
|
SCZ |
Human |
Astrocyte |
Increasing miR-223 |
Targeting glutamate receptors |
|
59
|
SCZ |
Human |
plasma |
Increasing miR-137 and decreasing COX6A2 |
Cortical microcircuit impairment |
|
60
|
SCZ |
Human |
Plasma |
Higher GFAP and lower α-II-spectrin concentrations |
Brain injury |
|
61
|
SCZ |
Human |
blood |
miR-137, miR-22-3p, miR-92A-3p |
A combination of these factors can be used as biomarkers of SZ |
|
62
|
Depression |
Human |
blood |
Higher miR-186-5p and lower SERPINF1 |
Suppression of SERPINF1 in the hippocampus by miR-186-5p |
|
63
|
Depression |
Human |
plasma |
Insulin receptor substrate -1 |
increase in patients |
|
64
|
Depression |
Human |
Serum |
higher miR-9-5p |
Increases microglia M1 polarization and exacerbation of depressive symptoms. |
|
65
|
Depression |
Human |
plasma |
miR-30d-5p, miR-486-5p, miR-21-5p |
changed during antidepressant treatment and related to antidepressant treatment response |
|
66
|
Depression |
Human |
blood |
Higher IL-34 |
neuron-related blood biomarkers |
|
66
|
Depression |
Human |
blood |
Synaptophysin and TNFR1 |
positively associated with severities of depression |
|
67
|
Depression |
Human |
plasma |
Lower mitofusin 2 and cyclophilin D |
neuron-related plasma biomarkers |
|
68
|
Depression |
Human |
Serum |
Upregulation of hsa-miR-139-5p |
upregulated in patients |
|
69
|
Depression and BD |
Human |
Blood |
Higher miR-132 |
Target CLOCK gene |
|
70
|
BD |
Human |
serum |
sugar metabolism dysfunction |
Molecular pathways underlying BP pathogenesis |
|
71
|
BD |
Human |
blood |
lower tryptophan, xanthurenic acid, kynurenine, and kynurenic acid |
Have potential for use as biomarkers |
|
72
|
BD |
Human |
plasma |
p-JNK, p-p38-MAPK, p-ERK1/2 |
higher in BD patients treated with infliximab |
|
73
|
BD |
Human |
serum |
phosphatidylinositol |
The level of phosphatidylinositol has an association with BD risk |
|
74
|
Autism |
Human |
blood |
Abnormal SVAT |
A marker for the development of neural circuits |
|
75
|
Autism |
Mice |
brain |
Higher IL-1 and proinflammatory cytokines |
Inflammation is a marker of autism |
|
76
|
Epilepsy |
Human |
Plasma |
hsa-miR-184 downregulation |
Modulate apoptosis, inflammation, and neuronal death after an epileptic state |
|
77
|
AD |
Human |
Plasma |
miR-190-5p, miR-223-3p, miR-100-3p |
showed a significant dysregulation |
|
78
|
AD |
Human |
plasma |
FGF, IGF-1, HGF |
significantly lower in AD patients |
Abbreviations: SCZ, schizophrenia; BD, bipolar disorder; AD, Alzheimer’s disease.
Role of EVs in MDD
Depression affects millions of people worldwide; it is one of the most prevalent mental disorders, and globally, the incidence is on the rise.79 In addition to a high recurrence rate and suicidal tendencies, patients also experience limited treatment effectiveness.80 Inflammatory hypotheses suggest that cytokines contribute to the pathophysiology of MDD and that changes in peripheral cytokine levels are associated with responses to antidepressants that include norepinephrine reuptake inhibitors (SNRIs), selective serotonin reuptake inhibitors (SSRIs), and serotonin.81 The etiology of MDD involves various mechanisms, including systemic immune activation, alterations in neurotransmitters and metabolism, dysfunction of the hypothalamic-pituitary-adrenal (HPA) axis, changes in the intestinal microbiome, and abnormalities in synaptic plasticity and mitochondrial function. Despite recent progress, no valid biomarkers have been detected to measure therapeutic response, disease state, or predict individual reactions to the therapeutic protocol.82 Due to the overlap between symptoms of MDD and other neuropsychiatric diseases, the diagnosis and prognosis of MDD require the detection of specific biomarkers.83 Studies targeting the role of EVs in MDD have provided important clues about potential mechanisms underlying this debilitating condition.84 Dysregulation of EV miRNA cargo has been observed among individuals with MDD, highlighting their involvement in regulating key molecular pathways relevant to depression.85 Furthermore, altered levels of EV-associated proteins involved in neuroplasticity and stress response have also been reported among MDD patients. The miRNA profile in neuron-derived plasma EVs of depressed patients showed the dysregulated miRNAs and their effect on postsynaptic density, cell growth, and axon formation, which can explain one of the reasons for a reduction in hippocampus volume in MDD patients.86 Microglia play a crucial role in MDD pathogenesis by inhibiting neurogenesis and the discharge of excitatory neurons through the secretion of EVs with higher levels of miR-146a-5p, which targets Krüppel-like factor 4 (KLF4). Additionally, EVs with higher miR-124 levels may enhance hippocampal neurogenesis and promote the polarization of M2 microglia.87
To maintain CNS function and structure, astrocytes play a critical role, and their EVs contain miRNAs that regulate adult neurogenesis and stress responses.88 In normal conditions, astrocytes release EVs that mediate TGF-β signal activation through fibulin-2, resulting in synapse formation.89 EVs with miR-200a-3p have a neuroprotective role by downregulating MKK4, while the existence of fibroblast growth factor-2 (FGF-2) helps to maintain BBB integrity.90 The innate immune system, particularly aberrant Toll-like receptor (TLR) expression, may play a role in MDD etiology. During the development of MDD, negative regulators of TLR signal pathways control the magnitude and duration of cytokine storms induced by TLRs.79 It has been found that plasma EVs deliver sigma-1 receptors to the CNS, alleviate inflammation, and improve depression-like behavior in animal models.91 Fang et al showed that chronic stress affects serum EVs microRNA content, leading to alterations in important signaling pathways, such as MAPK, Wnt, and mTOR, and impairing neuroplasticity and inflammation. This led to changes in the levels of neurotrophic factors (i.e., BDNF) and synaptic proteins in mood-regulating brain regions (e.g., the hippocampus and prefrontal cortex, or PFC). Altered microRNAs in EVs also disrupt neurogenesis and synaptic functions in these regions, providing insight into the depressive-like behaviors observed. Furthermore, these findings could have implications for EVs’ microRNAs as biomarkers for early detection and potential therapeutic targets for stress-induced depression.92 However, the study has limitations that should be noted, including that the study reported protein expression levels of important markers (BDNF, TrkB, synaptotagmin 1) only in the hippocampus and prefrontal cortex, and did not report depression-related changes in any other brain regions that may be important to consider. The study used a chronic unpredictable mild stress model in rats, and while a widely recognized behavioral model of depression, it may not reflect the entirety of human depression pathophysiology. Hung et al found the reduction of only miR-146a in serum EVs of MDD patients before antidepressant medication, and the levels of let-7e and miR-155 were increased following treatment with antidepressants. Compared to the remission group, the non-remission group exhibited significantly higher levels of let-7e, miR-21-5p, miR-223, and miR-146a, indicating that EVs negatively affect TLR4 signaling through the regulation of these microRNAs.79 The limitations of this study are the following: Variation in patient characteristics, severity of illness, medication regimens, and co-morbid conditions may bias miRNA expression and biomarker validity, and divergence in EV isolation methods may bias results by variability in miRNA extraction, extraction methods, quality control, and quantification. Wei et al found that hsa-miR-139-5p was significantly higher in MDD patients than in the control group. They isolated exosomes from the blood of MDD patients and injected them into normal mice. The results indicated that this group of mice showed depressive-like behaviors and had remarkably fewer DCX + cells in the subgranular zone, a marker of neurogenesis. Also, they isolated blood exosomes from healthy volunteers and injected them into mice models of depression that were subjected to chronic unpredictable mild stress. Compared to the control, this group of mice had a notably decreased total antioxidant capacity (T-AOC) and increased MDA levels, which are two markers for assessing oxidative stress.93 While the study identified EVs induced changes in oxidative stress and neurogenesis markers, neither the specific mechanisms nor the degree to which blood EVs enter the BBB to alter brain function could be determined.
It has been proposed that astrocytes are implicated in the pathophysiology of depression and could serve as a significant therapeutic target.94 These cells are the prime source of inflammatory cytokines.95 As a component of the TLR4 signaling pathway, leucine-rich repeats (Tril) are highly expressed in astrocytes and mediate pro-inflammatory changes by activating transcription factors.79 NK cell EV miR-207 may increase the release of monoamine factors in the brain by targeting Tril, thereby improving the behavior of mice and reducing the secretion of inflammatory factors.96 Ran et al conducted miRNA sequencing of serum EVs and identified 18 upregulated and 14 downregulated miRNAs in adolescents with childhood MDD. Of them, miR-450a-2-3p, miR-556-3p, and miR-2115-3p were remarkably different, especially miR-450a-2-3p. They also indicated the link between miR-450a-2-3p and trauma in childhood and MDD in adolescents.22 However, the study’s limitations include a relatively small sample size, which limits the generalizability and robustness of the findings. Additionally, the study did not examine the mechanistic underpinnings by which dysregulated miRNAs affect the pathophysiology of depression.
Significantly higher levels of microglial GLS1 mRNA, along with the neuroinflammation pathway, were reported by Ji et al in the prefrontal cortex of the depression model. Reduced neuroinflammation by GLS1-deficient microglia was a consequence of less reactive astrocytes, as GLS1 deficiency increases miR-7115-3p and miR-666-3p levels in EVs released from microglia, therefore suppressing astrocyte activation via blocking Serpina3n expression. These results highlight the promise that EVs hold as biomarkers that aid in comprehension, diagnosis, and the development of more effective treatments for MDD.97 A recent study elevated miR-182-5p levels as a biomarker and modulator of MDD, and the dysregulated miR-182-5p in EVs, known to play a role in depression through regulating critical genes related to HPA activity (e.g., FKBP5 and CRHR2), which control stress reactivity, will allow further examination of the EV-derived miR-182-5p in the potential for depression’s pathophysiology. The functional experiment of injecting MDD patient-derived EVs into the paraventricular nucleus (PVN) of mice demonstrated miR-182-5p-related HPA axis hyperactivity, as indicated by increased expression of corticotropin-releasing factor (CRF). Bioinformatics analysis also indicated pathways related to miR-182-5p, including protein transport, cytoskeletal regulation, and glutamatergic synapses, which relate to a depressive pathology. miR-182-5p within plasma sEVs contributes to MDD by modulating HPA axis dysfunction and synaptic changes, demonstrating the potential for vesicular miRNAs as biomarkers for diagnosis, treatment, and prevention of depression.98
In a recent paper, researchers reported that EVs derived from human umbilical cord mesenchymal stem cells (hUCMSCs) exhibit antidepressant effects by targeting specific pathophysiological pathways associated with depression. They found that these EVs produce a significant reduction in neuroinflammation by inhibiting pro-inflammatory cytokines, such as IL-1β and TNF-α, thereby normalizing the morphology of microglial activation in the hippocampus. Furthermore, as indicated by increased expression of BDNF, they promote neuronal survival and synaptic plasticity. EVs were also reported to stimulate neurogenesis and enhance the dendritic complexity of doublecortin-positive cells in the dentate gyrus, thereby restoring vulnerable interneuron connections after chronic stress. Moreover, their small size also promotes biodistribution and the ability to cross the blood-brain barrier, supporting immunomodulation in systemic circulation and the CNS. These various modulations of neuroimmune and neurotrophic systems by hucMSC-EVs contribute to the alleviation of depressive-like behaviors, making them a novel and efficacious intervention for treating depression.99
Role of EVs in AD
The ageing of the global population is leading to an annual increase in the number of AD patients and making it a major public health concern.100 A person with AD is most likely to be diagnosed once irreversible neuronal damage has occurred, as it is one of the conditions among the principal mortality causes without a preventative treatment or cure.101 AD symptoms mainly include neuropsychiatric abnormalities, progressive memory decline, and cognitive impairment.102 In AD pathogenesis, there is a silent accumulation of proteins, and symptoms do not occur until late pathological conditions.103 Individuals’ cognitive performance declines after this silent period, and memory and executive cognitive alterations first appear. The first molecular changes in AD pathogenesis can occur up to 15 years before symptoms appear.104 The most extensive study of EVs in the brain has been conducted on AD to date. The pathology of AD is confirmed by the presence of amyloid beta (Aβ) plaques and the formation of hyperphosphorylated tau (p-tau) tangles.105 The tau protein exists in several forms, including p-T181-tau, and p-S396-tau, and these molecules have been found in neuron EVs of AD patients compared to healthy controls.106 CSF-tau and CSF-Aβ can be used as diagnostic biomarkers for monitoring AD status.107 Studies have shown that with AD progression, the levels of EV Aβ42 protein are also increased.108 Other factors like growth-associated protein 43 (GAP43), neurogranin (NRGN), synaptosome-associated protein 25 (SNAP25), synaptotagmin, and repressor element 1-silencing transcription factor (REST) are statistically reduced.109 In a study by Kumar et al, the miRNA level and profile were found to be significantly different in plasma EVs of adults with mild cognitive impairment (MCI) and AD as compared to healthy individuals. They suggest downregulating miR-106b and miR-29a-5p, and overexpressing miR-106b-5p, miR-132-5p, and miR-106b-5p.110 The key limitations of the study are a moderate sample size of the population. Notably, this study cannot exclude potential confounders from peripheral sources of EVs, as blood EVs can originate from multiple organs and have been reported to differentiate in response to trauma or health states unrelated to brain pathology. In neuron-derived plasma EVs, the levels of miR-223-3p, miR-23a-3p, miR-190-5p, and miR-100-3p are between AD patients and controls (Figure 3). Along with these changes, the levels of FGF-2 and −13, hepatocyte growth factor (HGF), and type 1 insulin-like growth factor (IGF-1) were lowered in AD patients.111 Neurexin 2α, GluA4-containing glutamate receptor, neurogranin, and neuronal pentraxin 2 are proteins that originated from neuronal cells and are significantly decreased in the neuronal-derived plasma EVs of AD patients.112 According to a study by Koinuma et al, EV secretion affects both intracellular and extracellular Aβ levels. It also downregulates autophagosomes through siRNA. These findings imply that EVs are an important source of Aβ in the brain and that abnormal Aβ accumulation in EVs may play a role in the transition of age-dependent Aβ pathology.113
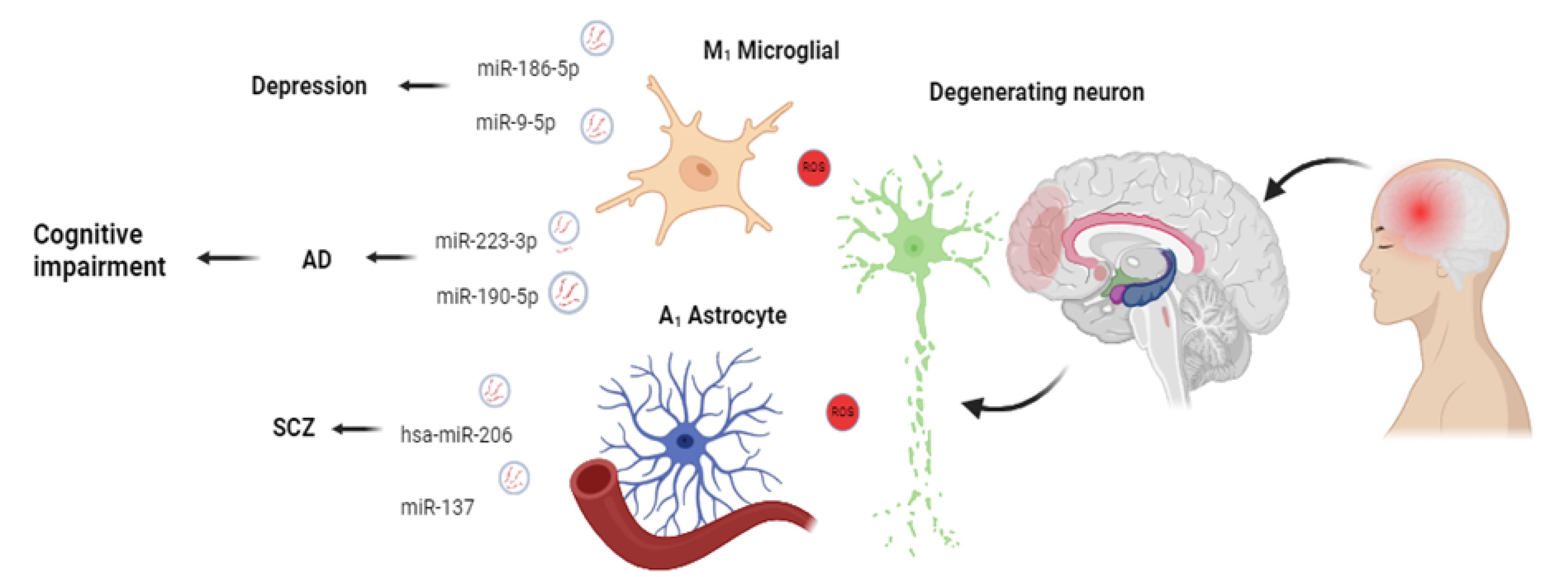
Figure 3.
The elevation of EV miR-186-5p is common in MDD. These features contribute to the suppression of SERPINF1 in the hippocampus. Besides, the increase of EV miR-9-5p promotes the polarization of microglia toward the M1 phenotype, resulting in the exacerbation of depressive symptoms. AD is characterized by significant dysregulation of EV miR-190-5p and miR-223-3p. Similar to AD and MDD, the levels of hsa-miR-206 (targeting BDNF mRNA), and miR-137 in EVs increase in the blood EV of SCZ patients. The alteration of genetic elements and various cargoes contributes to cognitive impairment and neuropsychiatric diseases
.
The elevation of EV miR-186-5p is common in MDD. These features contribute to the suppression of SERPINF1 in the hippocampus. Besides, the increase of EV miR-9-5p promotes the polarization of microglia toward the M1 phenotype, resulting in the exacerbation of depressive symptoms. AD is characterized by significant dysregulation of EV miR-190-5p and miR-223-3p. Similar to AD and MDD, the levels of hsa-miR-206 (targeting BDNF mRNA), and miR-137 in EVs increase in the blood EV of SCZ patients. The alteration of genetic elements and various cargoes contributes to cognitive impairment and neuropsychiatric diseases
Chen et al showed that cognitive deficits of AD, degradation of plaques, and brain glucose metabolism can be significantly improved by using mesenchymal stem cell (MSC)-derived EVs.114 Li et al found that bone marrow MSC (BMSC) EVs alleviated AD-like behaviors in a mouse model by increasing the expression of synapse-related proteins and BDNF, reducing the levels of IL-1β, IL-6, and TNF-α, and suppressing the hyperactivation of microglia and astrocytes.115 The limitations of this study are primarily related to the use of an AD animal model, which may not fully encompass all aspects of human AD pathology, thereby limiting clinical translation. The molecular mechanisms underlying the neuroprotective effects of BMSC-derived EVs, including their impact on BDNF pathways and neuroinflammation, require further exploration. High-throughput sequencing of EVs from hypoxia-pretreated adipose-derived stem cells (ADSCs) showed improved cognitive function. Based on the data, circ-Epc1 was an essential component of hypoxia-pretreated ADSC EVs with the potential to enhance mental function and microglia polarization from M1 to M2 type in AD mice.116 Li et al demonstrated that M2 microglia-derived EVs, when internalized by neurons, exhibited neuroprotective functions, increased cell viability, decreased Aβ plaque deposition, and improved the PINK1/Parkin pathway of autophagy.117 Iyaswamy et al presented engineering EVs that overexpress Fe65 and carried Corynoxine-B as an autophagy inducer. These EVs hijacked the interaction between Fe65 and APP, induced autophagy in amyloid precursor protein (APP)-expressing cells, and ameliorated the cognitive decline and pathogenesis in the AD model.118 The limitations of this study include the need to thoroughly assess the long-term safety of engineered EVs and any potential immune response triggered by their in vivo introduction into humans. Although the targeted delivery of engineered EV was beneficial, assessing off-target effects and biodistribution to other organs outside the brain, such as the liver and spleen, should always be conducted.
The application of EVs in clinical practice is still in the early stages, but advances in engineered exosomes for targeted drug delivery and purification technology could allow these findings to be applied.
The brain is comprised of approximately 86 billion neurons interconnected by trillions of synapses.119 This intricate communication network forms the basis of cognitive functions, emotions, and behavior. When an electrical impulse reaches the end of a neuron, it triggers the release of neurotransmitters into the synaptic space. These neurotransmitters bind to receptors on the adjacent neurons, generating a new electrical signal and continuing the transmission of information.120 In recent years, the increasing importance of exosomes in modulating synaptic communication has added a new dimension to our understanding of neuropsychiatric disorders.121 Aberrant exosome-mediated communication maycontribute to the synaptic dysfunction observed in conditions like MDD.122 In certain areas of the cerebral cortex and the hippocampus, interactions between the presynaptic proteins neurexin 2α (NRXN2α) and neuronal pentraxin 2 (NPTX2), along with their respective postsynaptic functional partners, the GluA4-containing glutamate receptor (AMPA4) and neuroligin 1 (NLGN1), enhance excitatory synaptic activity. Since the cognitive decline in AD participants is associated with early damage to these excitatory circuits in their brain tissues, levels of all proteins in plasma neuron-derived EVs, except for NPTX2, were significantly lower in the years before the onset of dementia, and levels of all proteins decreased significantly as dementia progressed.123 Moreover, the research by Cui et al provides evidence that EVs obtained from hypoxia-preconditioned mesenchymal stromal cells (MSCs) are capable of preventing memory deficits in AD by rescuing synaptic dysfunction and regulating inflammatory responses.124
Role of EVs in autism spectrum disorder
Autistic spectrum disorder (ASD) is a neurodevelopmental disorder, and it has been estimated that nearly 200,000 cases of autism occur per year.125 A large number of autistic patients suffer from intellectual disability and have lifelong developmental disabilities in adulthood due to severe blockages in neurological and mental development.126 Based on the statistics, ASD has a substantial economic and psychological impact on society and families.127 The symptoms of autism can generally be categorized into two main areas: social communication and repetitive behaviors. Children with autism may struggle to make eye contact, have difficulty understanding nonverbal cues, and have trouble initiating or maintaining conversations.128 There is no cure for autism, but early intervention and therapy can help improve outcomes for ASD children. Most therapies, for instance, special education, adjuvant drug therapy, and targeting intestinal microbes, cannot effectively improve the symptoms of autism patients.129 As SVAT-related RNA expression changes were detected in blood EVs of ASD patients, monitoring SVAT changes could be used to predict neural circuit development in the fetal brain.74 Researchers have found that EVs in the serum of ASD patients cause microglia to have more proinflammatory cytokines, particularly IL-1. In comparison with other sources of EVs, it is believed that umbilical cord MSCs can secrete EVs containing immune-suppressing factors, such as IL-10 and TGF-β, to inhibit pathological immune responses.28 MSC-EVs could alleviate repetitive behavioral phenotypes and have potency as cell-free, noninvasive therapies. They are also better clinical options due to their advantages in immunogenicity, clinical safety, ethics, and stability, as well as their simplicity in transportation and storage. A long-term improvement in the behavioral phenotype of autism was achieved by transplanting human bone marrow stem cells (BMSCs) into the lateral ventricles of BTBR mice, where EVs were the primary therapeutic mediators.75 The intranasal administration of human MSC EVs in rats with experimental perinatal brain damage reduced microglia-mediated neuroinflammation. This suggests that neuroinflammation, increased pro-inflammatory cytokine levels, and abnormal microglia/astrocyte responses play an important role in autism pathogenesis in autistic brains and animal models of autism.130
The results from a study by Qin et al show that engineered EVs bearing miR-137, which are modified with rabies virus glycoprotein (RVG) for targeting, ameliorate autism-like behaviors through neuroinflammation modulation and effects on brain glucose metabolism. MiR-137 could be increased in the brain after RVG-miR137-EVs treatment (the levels of miR-137 are decreased in autistic model mice and human patients), which lowers the activation of microglia and pro-inflammatory cytokine levels IL-1β and TNF-α. Mechanistically, the miR-137 targets and downregulates Toll-like receptor 4 (TLR4), thus inhibiting the TLR4/NF-κB signaling pathway. The downregulation of TLR4 decreases hypoxia-inducible factor 1α (HIF-1α) expression along with the essential glycolytic enzymes HK2, PKM2, and LDHA, resulting in decreased glycolysis and lactate function associated with the Warburg effect in activated microglia. Pharmacological inhibition of TLR4 or glycolysis, acting in parallel, decreases neuroinflammation and autism-like behaviors. In summary, miR-137–mediated repression of the TLR4/NF-κB/HIF-1α axis restores microglial metabolic and inflammatory behaviors, leading to improved neurological outcomes.131 This study needs further investigation of long-term effects, as behavioral tests were conducted at a single time point and may not fully capture the persistent therapeutic effects. The specific molecular mechanisms by which miR-137 modifies glucose metabolism and neuroinflammation through the TLR4/NF-κB and HIF-1α pathways also require further elucidation.
Table 2 provides a summary of EVs used for the treatment of various neuropsychiatric disorders.
Table 2.
Therapeutic Potential of EVs in Neuropsychiatric Disorders.
|
Reference
|
Diseases
|
Species
|
Source of EVs
|
Contents of EVs
|
Outcome
|
|
132
|
Depression |
Mice |
NK cells |
miR-207 overexpression |
Reduced inflammation and improved depressive symptoms |
|
133
|
Depression |
rats |
BMSC)-derived exosomes |
upregulating miR-26a |
Alleviate neuronal injury in the hippocampus |
|
134
|
Autism |
Mice |
umbilical cord-derived MSC |
IL-I0 and TGF-β |
Ameliorated symptoms of Autism |
|
114
|
AD |
Transgenic mice |
Wharton’s jelly MSCs |
HDAC4-targeting miRNA-29a |
Improve cognitive deficits and brain glucose metabolism |
|
116
|
AD |
Mice |
ADSCs |
circ-Epc1 |
shifted microglial M1/M2 polarization, and improved cognition |
|
117
|
AD |
Mice |
M2 Microglia |
miR-124-3p |
neuroprotective functions and improved PINK1/Parkin pathway of autophagy. |
Role of EVs in posttraumatic stress disorder (PTSD)
PTSD is one of the neuropsychiatric disorders that develops after experiencing a traumatic event, and its symptoms include intrusive thoughts, flashbacks, avoidance behaviors, and hyperarousal. Research indicated alterations in brain structure and function of patients with PTSD, such as a reduction in the volume of the hippocampus and abnormalities in the amygdala.135,136
The research on EVs in PTSD is still in its nascent stages, with a significant scarcity of large-scale studies compared to disorders like MDD or AD. Therefore, the current evidence is preliminary but points to several intriguing mechanisms. Kang et al demonstrated that changes in the expression of a set of circulating miRNAs associated with EVs, specifically related to the FK506-binding protein 5 (FKBP5) gene, are linked to PTSD.137 Lee et al also analyzed miRNAs in blood derived from PTSD patients and controlled by NGS and observed an increase in the concentration of miR-203a-3p. miR-203a-3p is associated with inflammatory pathways, serotonergic signaling, and neurotransmitter systems: various signaling pathways, cell cycle, inflammation, and metabolism.138 In a study, Bam et al found that miR-7113-5p is decreased in Peripheral Blood Mononuclear Cells (PBMCs) of PTSD patients. The researchers also showed that this downregulation of miR-7113-5p promotes an inflammatory response by targeting and upregulating the WNT signaling pathway.139 However, the sample size was small, and it is essential to replicate the study with larger and more diverse populations. Additionally, the role of dysregulated Wnt signaling as a driver of inflammation is supported; however, because the modulation of the Wnt pathway is context-dependent, we do not yet fully understand the downstream effects on the various immune cell subsets, nor the brain-immune interactions. Guo et al showed that EVs from PTSD subjects transport the chemokine CXCL8 through the protein CD63, which strengthens communication between astrocytes and neurons. These EVs exacerbated anxiety- and depression- like behavior in a PTSD mouse model, and even though silencing CXCL8, improved these symptoms. The authors conclude that the CD63-mediated delivery of CXCL8 via EVs is an important mechanism driving PTSD and a potential therapeutic target.140
Role of EVs in epilepsy
Epilepsy is characterized by abnormal discharges of neurons mostly in the hippocampus, amygdala, temporal, and frontal lobes,141 and the underlying reason for its onset is genetics, brain injuries (such as a stroke or a traumatic brain injury), developmental disorders, infections, and tumors. Diagnosis of this disorder is performed by evaluating the patient’s medical history, conducting a physical examination, performing electroencephalography (EEG), and utilizing brain imaging.142 In a study that characterized the effects of experimental status epilepticus on mouse hippocampal EVs and their miRNA content, miR-21-5p exhibited strong changes in CSF samples. miR-21-5p has been shown to regulate neurotrophin 3 mRNA and Mef2c transcripts, which encode neural transcription factors in the hippocampus. The efficacy of miR-21 in EVs may be associated with the status epilepticus or epileptogenesis. It has been shown that it may be useful in diagnosis.143 miR-346 and miR-331-3p were significantly downregulated in EVS from anterior brain cells in an animal model of epilepsy.144 Additionally, miR-21a-3p and miR-21a-5p, as well as miR-146a-5p, have been shown to be upregulated in animal models and humans.143 In addition to miR-194-2 and miR-15a, serum EVs in epilepsy patients were downregulated, and higher coagulation factor IX (F9) and lower thrombospondin-1 (TSP-1) were detected.145
Yu et al demonstrated that miR-106b-5p level increased and repulsive guidance molecule A (RGMa) and triggering receptor expressed on myeloid cell 2 (TREM2) levels decreased in the lithium-pilocarpine-induced Epilepsy mouse model. This finding indicated inflammation and microglial activation in the hippocampus of the Epilepsy mouse model.146 Mesenchymal stem cell EVs have been shown to have anti-inflammatory properties and restore activation of hippocampal astrocytes.147
Ballal et al demonstrated that exogenous GABA can be used to alleviate seizures in epilepsy when the GABA is carried and delivered by EVs derived from certain brain cells. EVs from GABAergic interneurons and EVs from medial ganglionic eminence cells reduced seizure frequency and severity in a rat model of epilepsy. Also, they improved depressive behavior and some memory functions. Cell-specific EV-mediated delivery of GABA provides a potentially exciting new treatment modality for the management of epilepsy.148 Nevertheless, these observations are limited due to rats with kainic acid-induced epilepsy being used in this study, and do not wholly represent the diversity, complexity, and multiple variants of human epilepsy. This study did not provide a rigorous evaluation of potential long-term effects and off-target effects with repeated dosing of EVs.
Role of EVs in bipolar disorder
BD is characterized by extreme mood changes that alternate between episodes of mania and depression, and Genetics and the environment both play a role in the development of this disease. During manic episodes, the symptoms include excessive energy, low need for sleep, and impulsivity. In contrast, during depressive episodes, patients experience sadness, hopelessness, low energy, and difficulty concentrating.149 The delay in diagnosis may lead to neuroprogression, which could be due to a cumulative effect of immune dysfunction, loss of neurotrophic factors, and increased oxidative stress.122 Research findings link EVs to the development and progression of BDs. Neuronal excitability in BP could be influenced by the EV-mediated transfer of miRNAs between neurons, which is dysregulated in patients.150
Ceylan et al analyzed the expression of miRNA in EVs of BD patients. The observation indicated significant downregulation of MiR-484, -652-3p, -142-3p, and remarkable upregulation of miR-185-5p. They performed bioinformatic analysis and identified several target pathways, including the PI3K/Akt signaling pathway, adhesion pathways, fatty acid biosynthesis/metabolism, and the extracellular matrix.151 Du et al performed ultraperformance liquid chromatography-tandem mass spectrometry (UPLC-MS-MS) and examined the serum-derived EVs of BD patients. They presented a total of 26 EV metabolites that exhibited differential expression in patients diagnosed with BD (n = 32) when compared to the control group. Furthermore, these differentially expressed metabolites were found to be significantly enriched in pathways associated with sugar metabolism.70 Study limitations include the use of a small sample size (32 BD patients and 40 healthy controls). Some confounders were not measured, such as smoking status, socioeconomic status, and the severity of the disease, which could affect the levels of the measured metabolites (Figure 4). The cross-sectional nature of this study does not permit firm conclusions about the changes in metabolites as the disease progresses.
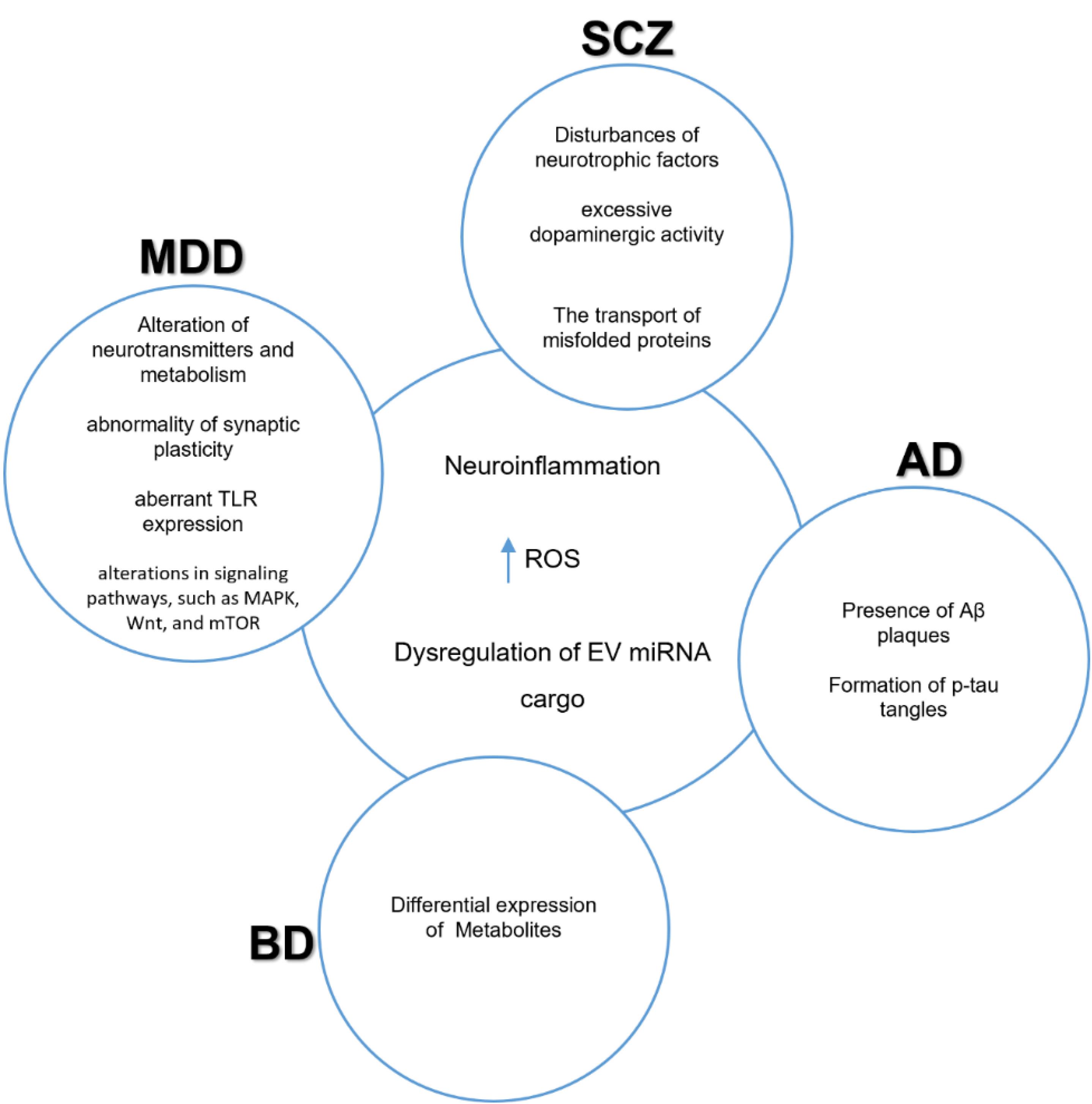
Figure 4.
Shared pathological mechanisms in neuropsychiatric disorders. schizophrenia (SCZ), major depressive disorder (MDD), Alzheimer’s disease (AD), and bipolar disorder (BD) have shared pathological mechanisms that include neuroinflammation, increased reactive oxygen species (ROS), and dysregulation of EV cargo composition151
.
Shared pathological mechanisms in neuropsychiatric disorders. schizophrenia (SCZ), major depressive disorder (MDD), Alzheimer’s disease (AD), and bipolar disorder (BD) have shared pathological mechanisms that include neuroinflammation, increased reactive oxygen species (ROS), and dysregulation of EV cargo composition151
Fries et al analyzed EV miRNAs in the blood of BD patients and controls, and their results revealed notable differences between the two groups of miRNAs, including miR-29c and miR-22. These miRNAs are involved in various signaling pathways in the brain, including serotonergic receptor type 5HT2 and netrin-mediated signaling pathway.152
Clinical translation
The clinical translation of engineered EVs presents numerous challenges, particularly in relation to regulatory approvals, reproducible manufacturing processes, and the design of well-structured clinical trials. A primary challenge concerning regulatory permissions involves the classification of EVs as drug products rather than as diagnostics. The critical regulatory issue pertains to establishing drug status, as the Food and Drug Administration (FDA) categorizes all EV-based therapeutics as biologics, which fall under the jurisdiction of its Center for Biologics Evaluation and Research. These products are required to adhere to the FDA’s stringent Investigational New Drug (IND) application process, necessitating comprehensive documentation about safety, efficacy, and purity. This process includes substantiating EV targeting and clearance through detailed biodistribution studies associated with the use of EVs. Moreover, it is imperative to document the immunogenicity and toxicity of the EVs, ensuring compliance with the minimum safety threshold established for regulated products. Given the biological heterogeneity of EV populations and their constituent materials, standardizing protocols for the production of engineered EVs in accordance with Good Manufacturing Practice (GMP) remains a significant challenge, particularly when compared to conventional therapeutic products. In addition to the complexities related to nanoparticles, efforts must be directed toward minimizing batch variability, scale-up manufacturing of EVs from bacterial, mammalian, or plant cell sources without compromising their integrity or potency, ensuring that all preparations are free from contaminants, and developing validated potency assays to confirm the therapeutic efficacy of the EVs. Furthermore, the selection of endpoints for clinical trials must be carefully tailored to address disease-specific requirements. This includes the analysis of EV biomarker levels, which may necessitate the incorporation of validated cognitive assessments for neuropsychiatric disorders such as MDD or BD. It is also essential to consider the impact of functional imaging techniques (e.g., fMRI or PET) on multi-modal assessments, which aim to encompass both the overall clinical response of symptoms and the biological mechanisms at play.153,154
Conclusion
The analysis of EVs in neuropsychiatric disorders enhances our understanding of their roles in gene expression and intercellular communication. This research underscores the potential of EVs as therapeutic targets and precise diagnostic tools. However, these studies have not yet transitioned into clinical applications due to several limitations, including the limited availability of disease-specific EV-based biomarkers, ambiguity concerning the precise functions of EVs, inconsistent research outcomes, small sample sizes, a lack of standardized detection methodologies, and concerns regarding the reliability of EVs. Recent investigations using proton magnetic resonance spectroscopy and high-throughput sequencing have aimed to identify potential biomarkers that can distinguish between various neuropsychiatric conditions. In contrast to invasive procedures such as tissue biopsies and lumbar punctures, EVs present numerous advantages in clinical practice, including the ability to reflect brain pathophysiology without requiring surgical interventions, thereby enhancing prognosis and facilitating the monitoring of disease progression. Consequently, there is a need for the development of methods based on the hallmarks of EVs associated with the early stages of neuropsychiatric disorders, which would allow for non-invasive, cost-effective, population-wide screening. Despite the lack of clear pathological features in neuropsychiatric diseases, omics exploration has revealed new potential protein and miRNA biomarkers that may assist in predicting disease status. Ongoing research in this domain has the potential to elucidate the underlying mechanisms of these disorders and contribute to the development of effective diagnostic and therapeutic tools. The capacity of EVs to deliver disease-specific biomarkers while providing therapeutic cargo signifies their promise as combined theragnostic platforms. Future studies must focus not only on the independent development of diagnostics and therapies but also on integrating both elements to create personalized diagnostic and treatment approaches for neuropsychiatric conditions. This review aims to summarize the advancements in the application of EVs as biomarkers and treatment modalities for neuropsychiatric disorders, which carry significant implications for clinical utilization.
Competing Interests
The authors declare that they have no conflicts of interest.
Ethical Approval
Not applicable.
References
- Andreassen OA, Hindley GF, Frei O, Smeland OB. New insights from the last decade of research in psychiatric genetics: discoveries, challenges and clinical implications. World Psychiatry 2023; 22(1):4-24. doi: 10.1002/wps.21034 [Crossref] [ Google Scholar]
- Nestler EJ, Hyman SE. Animal models of neuropsychiatric disorders. Nat Neurosci 2010; 13(10):1161-9. doi: 10.1038/nn.2647 [Crossref] [ Google Scholar]
- Si Q, Wu L, Pang D, Jiang P. Exosomes in brain diseases: pathogenesis and therapeutic targets. MedComm (2020) 2023; 4(3):e287. doi: 10.1002/mco2.287 [Crossref] [ Google Scholar]
- Comer AL, Carrier M, Tremblay M, Cruz-Martín A. The inflamed brain in schizophrenia: the convergence of genetic and environmental risk factors that lead to uncontrolled neuroinflammation. Front Cell Neurosci 2020; 14:274. doi: 10.3389/fncel.2020.00274 [Crossref] [ Google Scholar]
- Zhuo CJ, Hou WH, Jiang DG, Tian HJ, Wang LN, Jia F. Circular RNAs in early brain development and their influence and clinical significance in neuropsychiatric disorders. Neural Regen Res 2020; 15(5):817-23. doi: 10.4103/1673-5374.268969 [Crossref] [ Google Scholar]
- García-Gutiérrez MS, Navarrete F, Sala F, Gasparyan A, Austrich-Olivares A, Manzanares J. Biomarkers in psychiatry: concept, definition, types and relevance to the clinical reality. Front Psychiatry 2020; 11:432. doi: 10.3389/fpsyt.2020.00432 [Crossref] [ Google Scholar]
- Goričar K, Dolžan V, Lenassi M. Extracellular vesicles: a novel tool facilitating personalized medicine and pharmacogenomics in oncology. Front Pharmacol 2021; 12:671298. doi: 10.3389/fphar.2021.671298 [Crossref] [ Google Scholar]
- Wang R, Wang X, Zhang Y, Zhao H, Cui J, Li J. Emerging prospects of extracellular vesicles for brain disease theranostics. J Control Release 2022; 341:844-68. doi: 10.1016/j.jconrel.2021.12.024 [Crossref] [ Google Scholar]
- Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science 2020; 367(6478):eaau6977. doi: 10.1126/science.aau6977 [Crossref] [ Google Scholar]
- Nasiry D, Khalatbary AR. Stem cell-derived extracellular vesicle-based therapy for nerve injury: a review of the molecular mechanisms. World Neurosurg X 2023; 19:100201. doi: 10.1016/j.wnsx.2023.100201 [Crossref] [ Google Scholar]
- Saeedi S, Israel S, Nagy C, Turecki G. The emerging role of exosomes in mental disorders. Transl Psychiatry 2019; 9(1):122. doi: 10.1038/s41398-019-0459-9 [Crossref] [ Google Scholar]
- Schnatz A, Müller C, Brahmer A, Krämer-Albers EM. Extracellular vesicles in neural cell interaction and CNS homeostasis. FASEB Bioadv 2021; 3(8):577-92. doi: 10.1096/fba.2021-00035 [Crossref] [ Google Scholar]
- Zhao S, Sheng S, Wang Y, Ding L, Xu X, Xia X. Astrocyte-derived extracellular vesicles: a double-edged sword in central nervous system disorders. NeurosciBiobehav Rev 2021; 125:148-59. doi: 10.1016/j.neubiorev.2021.02.027 [Crossref] [ Google Scholar]
- Heidarzadeh M, Gürsoy-Özdemir Y, Kaya M, Eslami Abriz A, Zarebkohan A, Rahbarghazi R. Exosomal delivery of therapeutic modulators through the blood-brain barrier; promise and pitfalls. Cell Biosci 2021; 11(1):142. doi: 10.1186/s13578-021-00650-0 [Crossref] [ Google Scholar]
- Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signal 2021; 19(1):47. doi: 10.1186/s12964-021-00730-1 [Crossref] [ Google Scholar]
- Li H, Yuan Y, Xie Q, Dong Z. Exosomes: potential targets for the diagnosis and treatment of neuropsychiatric disorders. J Transl Med 2024; 22(1):115. doi: 10.1186/s12967-024-04893-6 [Crossref] [ Google Scholar]
- Bahram Sangani N, Gomes AR, Curfs LM, Reutelingsperger CP. The role of extracellular vesicles during CNS development. Prog Neurobiol 2021; 205:102124. doi: 10.1016/j.pneurobio.2021.102124 [Crossref] [ Google Scholar]
- Verderio C, Gabrielli M, Giussani P. Role of sphingolipids in the biogenesis and biological activity of extracellular vesicles. J Lipid Res 2018; 59(8):1325-40. doi: 10.1194/jlr.R083915 [Crossref] [ Google Scholar]
- Krylova SV, Feng D. The machinery of exosomes: biogenesis, release, and uptake. Int J Mol Sci 2023; 24(2):1337. doi: 10.3390/ijms24021337 [Crossref] [ Google Scholar]
- Joshi BS, de Beer MA, Giepmans BN, Zuhorn IS. Endocytosis of extracellular vesicles and release of their cargo from endosomes. ACS Nano 2020; 14(4):4444-55. doi: 10.1021/acsnano.9b10033 [Crossref] [ Google Scholar]
- Babuta M, Szabo G. Extracellular vesicles in inflammation: focus on the microRNA cargo of EVs in modulation of liver diseases. J Leukoc Biol 2022; 111(1):75-92. doi: 10.1002/jlb.3mir0321-156r [Crossref] [ Google Scholar]
- Ran LY, Kong YT, Xiang JJ, Zeng Q, Zhang CY, Shi L. Serum extracellular vesicle microRNA dysregulation and childhood trauma in adolescents with major depressive disorder. Bosn J Basic Med Sci 2022; 22(6):959-71. doi: 10.17305/bjbms.2022.7110 [Crossref] [ Google Scholar]
- Silvestro S, Bramanti P, Mazzon E. Role of miRNAs in Alzheimer’s disease and possible fields of application. Int J Mol Sci 2019; 20(16):3979. doi: 10.3390/ijms20163979 [Crossref] [ Google Scholar]
- Luo D, Liu H, Liu H, Wu W, Zhu H, Ge W. Long RNA profiles of human brain extracellular vesicles provide new insights into the pathogenesis of Alzheimer’s disease. Aging Dis 2023; 14(1):229-44. doi: 10.14336/ad.2022.0607 [Crossref] [ Google Scholar]
- Mattick JS, Amaral PP, Carninci P, Carpenter S, Chang HY, Chen LL. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nat Rev Mol Cell Biol 2023; 24(6):430-47. doi: 10.1038/s41580-022-00566-8 [Crossref] [ Google Scholar]
- Zhou WY, Cai ZR, Liu J, Wang DS, Ju HQ, Xu RH. Circular RNA: metabolism, functions and interactions with proteins. Mol Cancer 2020; 19(1):172. doi: 10.1186/s12943-020-01286-3 [Crossref] [ Google Scholar]
- Shao Y, Jiang Y. Circular RNAs in toxicology. Toxicol Sci 2021; 179(2):149-61. doi: 10.1093/toxsci/kfaa173 [Crossref] [ Google Scholar]
- Gomes AR, Bahram Sangani N, Fernandes TG, Diogo MM, Curfs LM, Reutelingsperger CP. Extracellular vesicles in CNS developmental disorders. Int J Mol Sci 2020; 21(24):9428. doi: 10.3390/ijms21249428 [Crossref] [ Google Scholar]
- Hornung S, Dutta S, Bitan G. CNS-derived blood exosomes as a promising source of biomarkers: opportunities and challenges. Front Mol Neurosci 2020; 13:38. doi: 10.3389/fnmol.2020.00038 [Crossref] [ Google Scholar]
- Chen T, Chen D, Su W, Liang J, Liu X, Cai M. Extracellular vesicles as vital players in drug delivery: a focus on clinical disease treatment. Front BioengBiotechnol 2025; 13:1600227. doi: 10.3389/fbioe.2025.1600227 [Crossref] [ Google Scholar]
- Stranford DM, Simons LM, Berman KE, Cheng L, DiBiase BN, Hung ME. Genetically encoding multiple functionalities into extracellular vesicles for the targeted delivery of biologics to T cells. Nat Biomed Eng 2024; 8(4):397-414. doi: 10.1038/s41551-023-01142-x [Crossref] [ Google Scholar]
- Walker S, Busatto S, Pham A, Tian M, Suh A, Carson K. Extracellular vesicle-based drug delivery systems for cancer treatment. Theranostics 2019; 9(26):8001-17. doi: 10.7150/thno.37097 [Crossref] [ Google Scholar]
- Yang Y, Wang Y, Wei S, Zhou C, Yu J, Wang G. Extracellular vesicles isolated by size-exclusion chromatography present suitability for RNomics analysis in plasma. J Transl Med 2021; 19(1):104. doi: 10.1186/s12967-021-02775-9 [Crossref] [ Google Scholar]
- Williams S, Fernandez-Rhodes M, Law A, Peacock B, Lewis MP, Davies OG. Comparison of extracellular vesicle isolation processes for therapeutic applications. J Tissue Eng 2023; 14:20417314231174609. doi: 10.1177/20417314231174609 [Crossref] [ Google Scholar]
- Lang JB, Buck MC, Rivière J, Stambouli O, Sachenbacher K, Choudhary P. Comparative analysis of extracellular vesicle isolation methods from human AML bone marrow cells and AML cell lines. Front Oncol 2022; 12:949261. doi: 10.3389/fonc.2022.949261 [Crossref] [ Google Scholar]
- Suresh PS, Zhang Q. Comprehensive comparison of methods for isolation of extracellular vesicles from human plasma. J Proteome Res 2025; 24(6):2956-67. doi: 10.1021/acs.jproteome.5c00149 [Crossref] [ Google Scholar]
- Veerman RE, Teeuwen L, Czarnewski P, Güclüler Akpinar G, Sandberg A, Cao X. Molecular evaluation of five different isolation methods for extracellular vesicles reveals different clinical applicability and subcellular origin. J Extracell Vesicles 2021; 10(9):e12128. doi: 10.1002/jev2.12128 [Crossref] [ Google Scholar]
- Figueroa-Hall LK, Burrows K, Alarbi AM, Hannafon BN, Hladik C, Tan C. Comparison of methods for isolation and characterization of total and astrocyte-enriched extracellular vesicles from human serum and plasma. J Extracell Biol 2025; 4(2):e70035. doi: 10.1002/jex2.70035 [Crossref] [ Google Scholar]
- Brennan K, Martin K, FitzGerald SP, O’Sullivan J, Wu Y, Blanco A. A comparison of methods for the isolation and separation of extracellular vesicles from protein and lipid particles in human serum. Sci Rep 2020; 10(1):1039. doi: 10.1038/s41598-020-57497-7 [Crossref] [ Google Scholar]
- Du X, Lv J, Feng J, Li X, Gao Y, Wang X. Plasma exosomes lncRNA-miRNA-mRNA network construction and its diagnostic efficacy identification in first-episode schizophrenia. BMC Psychiatry 2023; 23(1):611. doi: 10.1186/s12888-023-05052-9 [Crossref] [ Google Scholar]
- Harvey PD, Bosia M, Cavallaro R, Howes OD, Kahn RS, Leucht S. Cognitive dysfunction in schizophrenia: an expert group paper on the current state of the art. Schizophr Res Cogn 2022; 29:100249. doi: 10.1016/j.scog.2022.100249 [Crossref] [ Google Scholar]
- Baird A, Rathod S, Hansen L, Appleby L, Rodway C, Turnbull P. Suicide and psychosis: comparing the characteristics of patients who died by suicide following recent onset and longer duration of schizophrenia and other primary psychotic disorders, 2008-2021. Schizophr Bull 2025:sbaf009. doi: 10.1093/schbul/sbaf009.
- Frydecka D, Krzystek-Korpacka M, Lubeiro A, Stramecki F, Stańczykiewicz B, Beszłej JA. Profiling inflammatory signatures of schizophrenia: a cross-sectional and meta-analysis study. Brain BehavImmun 2018; 71:28-36. doi: 10.1016/j.bbi.2018.05.002 [Crossref] [ Google Scholar]
- Borgmann-Winter KE, Wang K, Bandyopadhyay S, Doostparast Torshizi A, Blair IA, Hahn CG. The proteome and its dynamics: a missing piece for integrative multi-omics in schizophrenia. Schizophr Res 2020; 217:148-61. doi: 10.1016/j.schres.2019.07.025 [Crossref] [ Google Scholar]
- Kurtulmuş A, Koçana C, Toprak SF, Sözer S. The role of Extracellular Genomic Materials (EGMs) in psychiatric disorders. Transl Psychiatry 2023; 13(1):262. doi: 10.1038/s41398-023-02549-5 [Crossref] [ Google Scholar]
- Oraki Kohshour M, Papiol S, Delalle I, Rossner MJ, Schulze TG. Extracellular vesicle approach to major psychiatric disorders. Eur Arch Psychiatry Clin Neurosci 2023; 273(6):1279-93. doi: 10.1007/s00406-022-01497-3 [Crossref] [ Google Scholar]
- Cariaga-Martinez A, Saiz-Ruiz J, Alelú-Paz R. From linkage studies to epigenetics: what we know and what we need to know in the neurobiology of schizophrenia. Front Neurosci 2016; 10:202. doi: 10.3389/fnins.2016.00202 [Crossref] [ Google Scholar]
- Du Y, Yu Y, Hu Y, Li XW, Wei ZX, Pan RY. Genome-wide, integrative analysis implicates exosome-derived microRNA dysregulation in schizophrenia. Schizophr Bull 2019; 45(6):1257-66. doi: 10.1093/schbul/sby191 [Crossref] [ Google Scholar]
- McCutcheon RA, Abi-Dargham A, Howes OD. Schizophrenia, dopamine and the striatum: from biology to symptoms. Trends Neurosci 2019; 42(3):205-20. doi: 10.1016/j.tins.2018.12.004 [Crossref] [ Google Scholar]
- Funahashi Y, Yoshino Y, Iga JI, Ueno SI. Impact of clozapine on the expression of miR-675-3p in plasma exosomes derived from patients with schizophrenia. World J Biol Psychiatry 2023; 24(4):303-13. doi: 10.1080/15622975.2022.2104924 [Crossref] [ Google Scholar]
- Du Y, Chen L, Li XS, Li XL, Xu XD, Tai SB. Metabolomic identification of exosome-derived biomarkers for schizophrenia: a large multicenter study. Schizophr Bull 2021; 47(3):615-23. doi: 10.1093/schbul/sbaa166 [Crossref] [ Google Scholar]
- Goetzl EJ, Srihari VH, Guloksuz S, Ferrara M, Tek C, Heninger GR. Decreased mitochondrial electron transport proteins and increased complement mediators in plasma neural-derived exosomes of early psychosis. Transl Psychiatry 2020; 10(1):361. doi: 10.1038/s41398-020-01046-3 [Crossref] [ Google Scholar]
- Guo C, Li J, Guo M, Bai R, Lei G, Sun H. Aberrant expressions of MIAT and PVT1 in serum exosomes of schizophrenia patients. Schizophr Res 2022; 240:71-2. doi: 10.1016/j.schres.2021.12.013 [Crossref] [ Google Scholar]
- Rashidi SK, Kalirad A, Rafie S, Behzad E, Ansari Dezfouli M. The role of microRNAs in neurobiology and pathophysiology of the hippocampus. Front Mol Neurosci 2023; 16:1226413. doi: 10.3389/fnmol.2023.1226413 [Crossref] [ Google Scholar]
- Hurşitoğlu O, Orhan F, Belge Kurutaş E, Doğaner A, Durmuş HT, Kopar H. Diagnostic performance of increased malondialdehyde level and oxidative stress in patients with schizophrenia. Noro Psikiyatr Ars 2021; 58(3):184-8. doi: 10.29399/npa.27372 [Crossref] [ Google Scholar]
- Peng H, Ouyang L, Li D, Li Z, Yuan L, Fan L. Short-chain fatty acids in patients with schizophrenia and ultra-high risk population. Front Psychiatry 2022; 13:977538. doi: 10.3389/fpsyt.2022.977538 [Crossref] [ Google Scholar]
- Lee EE, Winston-Gray C, Barlow JW, Rissman RA, Jeste DV. Plasma levels of neuron-and astrocyte-derived exosomal amyloid beta1-42, amyloid beta1-40, and phosphorylated tau levels in schizophrenia patients and non-psychiatric comparison subjects: relationships with cognitive functioning and psychopathology. Front Psychiatry 2020; 11:532624. doi: 10.3389/fpsyt.2020.532624 [Crossref] [ Google Scholar]
- Amoah SK, Rodriguez BA, Logothetis CN, Chander P, Sellgren CM, Weick JP. Exosomal secretion of a psychosis-altered miRNA that regulates glutamate receptor expression is affected by antipsychotics. Neuropsychopharmacology 2020; 45(4):656-65. doi: 10.1038/s41386-019-0579-1 [Crossref] [ Google Scholar]
- Khadimallah I, Jenni R, Cabungcal JH, Cleusix M, Fournier M, Beard E. Mitochondrial, exosomal miR137-COX6A2 and gamma synchrony as biomarkers of parvalbumin interneurons, psychopathology, and neurocognition in schizophrenia. Mol Psychiatry 2022; 27(2):1192-204. doi: 10.1038/s41380-021-01313-9 [Crossref] [ Google Scholar]
- Ranganathan M, Rahman M, Ganesh S, D’Souza DC, Skosnik PD, Radhakrishnan R. Analysis of circulating exosomes reveals a peripheral signature of astrocytic pathology in schizophrenia. World J Biol Psychiatry 2022; 23(1):33-45. doi: 10.1080/15622975.2021.1907720 [Crossref] [ Google Scholar]
- Ma J, Shang S, Wang J, Zhang T, Nie F, Song X. Identification of miR-22-3p, miR-92a-3p, and miR-137 in peripheral blood as biomarker for schizophrenia. Psychiatry Res 2018; 265:70-6. doi: 10.1016/j.psychres.2018.03.080 [Crossref] [ Google Scholar]
- Jiang M, Gu YF, Cai JF, Wang A, He Y, Feng YL. MiR-186-5p dysregulation leads to depression-like behavior by de-repressing SERPINF1 in hippocampus. Neuroscience 2021; 479:48-59. doi: 10.1016/j.neuroscience.2021.10.005 [Crossref] [ Google Scholar]
- Nasca C, Dobbin J, Bigio B, Watson K, de Angelis P, Kautz M. Insulin receptor substrate in brain-enriched exosomes in subjects with major depression: on the path of creation of biosignatures of central insulin resistance. Mol Psychiatry 2021; 26(9):5140-9. doi: 10.1038/s41380-020-0804-7 [Crossref] [ Google Scholar]
- Xian X, Cai LL, Li Y, Wang RC, Xu YH, Chen YJ. Neuron secrete exosomes containing miR-9-5p to promote polarization of M1 microglia in depression. J Nanobiotechnology 2022; 20(1):122. doi: 10.1186/s12951-022-01332-w [Crossref] [ Google Scholar]
- Saeedi S, Nagy C, Ibrahim P, Théroux JF, Wakid M, Fiori LM. Neuron-derived extracellular vesicles enriched from plasma show altered size and miRNA cargo as a function of antidepressant drug response. Mol Psychiatry 2021; 26(12):7417-24. doi: 10.1038/s41380-021-01255-2 [Crossref] [ Google Scholar]
- Kuwano N, Kato TA, Mitsuhashi M, Sato-Kasai M, Shimokawa N, Hayakawa K. Neuron-related blood inflammatory markers as an objective evaluation tool for major depressive disorder: an exploratory pilot case-control study. J Affect Disord 2018; 240:88-98. doi: 10.1016/j.jad.2018.07.040 [Crossref] [ Google Scholar]
- Goetzl EJ, Wolkowitz OM, Srihari VH, Reus VI, Goetzl L, Kapogiannis D. Abnormal levels of mitochondrial proteins in plasma neuronal extracellular vesicles in major depressive disorder. Mol Psychiatry 2021; 26(12):7355-62. doi: 10.1038/s41380-021-01268-x [Crossref] [ Google Scholar]
- Kouter K, Paska AV. Biomarkers for suicidal behavior: miRNAs and their potential for diagnostics through liquid biopsy - a systematic review. Epigenomics 2020; 12(24):2219-35. doi: 10.2217/epi-2020-0196 [Crossref] [ Google Scholar]
- Gruzdev SK, Yakovlev AA, Druzhkova TA, Guekht AB, Gulyaeva NV. The missing link: how exosomes and miRNAs can help in bridging psychiatry and molecular biology in the context of depression, bipolar disorder and schizophrenia. Cell Mol Neurobiol 2019; 39(6):729-50. doi: 10.1007/s10571-019-00684-6 [Crossref] [ Google Scholar]
- Du Y, Dong JH, Chen L, Liu H, Zheng GE, Chen GY. Metabolomic identification of serum exosome-derived biomarkers for bipolar disorder. Oxid Med Cell Longev 2022; 2022:5717445. doi: 10.1155/2022/5717445 [Crossref] [ Google Scholar]
- Almulla AF, Thipakorn Y, Vasupanrajit A, Abo Algon AA, Tunvirachaisakul C, Hashim Aljanabi AA. The tryptophan catabolite or kynurenine pathway in major depressive and bipolar disorder: a systematic review and meta-analysis. Brain BehavImmun Health 2022; 26:100537. doi: 10.1016/j.bbih.2022.100537 [Crossref] [ Google Scholar]
- Mansur RB, Delgado-Peraza F, Subramaniapillai M, Lee Y, Iacobucci M, Nasri F. Exploring brain insulin resistance in adults with bipolar depression using extracellular vesicles of neuronal origin. J Psychiatr Res 2021; 133:82-92. doi: 10.1016/j.jpsychires.2020.12.007 [Crossref] [ Google Scholar]
- Knowles EE, Meikle PJ, Huynh K, Göring HH, Olvera RL, Mathias SR. Serum phosphatidylinositol as a biomarker for bipolar disorder liability. Bipolar Disord 2017; 19(2):107-15. doi: 10.1111/bdi.12468 [Crossref] [ Google Scholar]
- Fang Y, Wan C, Wen Y, Wu Z, Pan J, Zhong M. Autism-associated synaptic vesicle transcripts are differentially expressed in maternal plasma exosomes of physiopathologic pregnancies. J Transl Med 2021; 19(1):154. doi: 10.1186/s12967-021-02821-6 [Crossref] [ Google Scholar]
- Alessio N, Brigida AL, Peluso G, Antonucci N, Galderisi U, Siniscalco D. Stem cell-derived exosomes in autism spectrum disorder. Int J Environ Res Public Health 2020; 17(3):944. doi: 10.3390/ijerph17030944 [Crossref] [ Google Scholar]
- Huang LG, Luo YH, Xu JW, Lu QC. Plasma Exosomal miRNAs expression profile in mesial temporal lobe epilepsy with hippocampal sclerosis: case-control study and analysis of potential functions. Front Mol Neurosci 2020; 13:584828. doi: 10.3389/fnmol.2020.584828 [Crossref] [ Google Scholar]
- Serpente M, Fenoglio C, D’Anca M, Arcaro M, Sorrentino F, Visconte C. MiRNA profiling in plasma neural-derived small extracellular vesicles from patients with Alzheimer’s disease. Cells 2020; 9(6):1443. doi: 10.3390/cells9061443 [Crossref] [ Google Scholar]
- Goetzl EJ, Nogueras-Ortiz C, Mustapic M, Mullins RJ, Abner EL, Schwartz JB. Deficient neurotrophic factors of CSPG4-type neural cell exosomes in Alzheimer disease. FASEB J 2019; 33(1):231-8. doi: 10.1096/fj.201801001 [Crossref] [ Google Scholar]
- Hung YY, Chou CK, Yang YC, Fu HC, Loh EW, Kang HY. Exosomal let-7e, miR-21-5p, miR-145, miR-146a and miR-155 in predicting antidepressants response in patients with major depressive disorder. Biomedicines 2021; 9(10):1428. doi: 10.3390/biomedicines9101428 [Crossref] [ Google Scholar]
- Zhang B, Fu W, Guo Y, Chen Y, Jiang C, Li X. Effectiveness of mindfulness-based cognitive therapy against suicidal ideation in patients with depression: a systematic review and meta-analysis. J Affect Disord 2022; 319:655-62. doi: 10.1016/j.jad.2022.09.091 [Crossref] [ Google Scholar]
- Liu JJ, Wei YB, Strawbridge R, Bao Y, Chang S, Shi L. Peripheral cytokine levels and response to antidepressant treatment in depression: a systematic review and meta-analysis. Mol Psychiatry 2020; 25(2):339-50. doi: 10.1038/s41380-019-0474-5 [Crossref] [ Google Scholar]
- Thase ME. Using biomarkers to predict treatment response in major depressive disorder: evidence from past and present studies. Dialogues Clin Neurosci 2014; 16(4):539-44. doi: 10.31887/DCNS.2014.16.4/mthase [Crossref] [ Google Scholar]
- Yang T, Frangou S, Lam RW, Huang J, Su Y, Zhao G. Probing the clinical and brain structural boundaries of bipolar and major depressive disorder. Transl Psychiatry 2021; 11(1):48. doi: 10.1038/s41398-020-01169-7 [Crossref] [ Google Scholar]
- Honorato-Mauer J, Xavier G, Ota VK, Chehimi SN, Mafra F, Cuóco C. Alterations in microRNA of extracellular vesicles associated with major depression, attention-deficit/hyperactivity and anxiety disorders in adolescents. Transl Psychiatry 2023; 13(1):47. doi: 10.1038/s41398-023-02326-4 [Crossref] [ Google Scholar]
- Cuomo-Haymour N, Kaiser S, Hartmann-Riemer M, Guetter K, Klaus F, Cathomas F. Differential expression of serum extracellular vesicle microRNAs and analysis of target-gene pathways in major depressive disorder. Biomark Neuropsychiatry 2022; 6:100049. doi: 10.1016/j.bionps.2022.100049 [Crossref] [ Google Scholar]
- Li LD, Naveed M, Du ZW, Ding H, Gu K, Wei LL. Abnormal expression profile of plasma-derived exosomal microRNAs in patients with treatment-resistant depression. Hum Genomics 2021; 15(1):55. doi: 10.1186/s40246-021-00354-z [Crossref] [ Google Scholar]
- Fan C, Li Y, Lan T, Wang W, Long Y, Yu SY. Microglia secrete miR-146a-5p-containing exosomes to regulate neurogenesis in depression. Mol Ther 2022; 30(3):1300-14. doi: 10.1016/j.ymthe.2021.11.006 [Crossref] [ Google Scholar]
- Upadhya R, Zingg W, Shetty S, Shetty AK. Astrocyte-derived extracellular vesicles: neuroreparative properties and role in the pathogenesis of neurodegenerative disorders. J Control Release 2020; 323:225-39. doi: 10.1016/j.jconrel.2020.04.017 [Crossref] [ Google Scholar]
- Patel MR, Weaver AM. Astrocyte-derived small extracellular vesicles promote synapse formation via fibulin-2-mediated TGF-β signaling. Cell Rep 2021; 34(10):108829. doi: 10.1016/j.celrep.2021.108829 [Crossref] [ Google Scholar]
- Shakespear N, Ogura M, Yamaki J, Homma Y. Astrocyte-derived exosomal microRNA miR-200a-3p prevents MPP( + )-induced apoptotic cell death through down-regulation of MKK4. Neurochem Res 2020; 45(5):1020-33. doi: 10.1007/s11064-020-02977-5 [Crossref] [ Google Scholar]
- Wang Y, Gao C, Gao T, Zhao L, Zhu S, Guo L. Plasma exosomes from depression ameliorate inflammation-induced depressive-like behaviors via sigma-1 receptor delivery. Brain BehavImmun 2021; 94:225-34. doi: 10.1016/j.bbi.2021.02.004 [Crossref] [ Google Scholar]
- Fang K, Xu JX, Chen XX, Gao XR, Huang LL, Du AQ. Differential serum exosome microRNA profile in a stress-induced depression rat model. J Affect Disord 2020; 274:144-58. doi: 10.1016/j.jad.2020.05.017 [Crossref] [ Google Scholar]
- Wei ZX, Xie GJ, Mao X, Zou XP, Liao YJ, Liu QS. Exosomes from patients with major depression cause depressive-like behaviors in mice with involvement of miR-139-5p-regulated neurogenesis. Neuropsychopharmacology 2020; 45(6):1050-8. doi: 10.1038/s41386-020-0622-2 [Crossref] [ Google Scholar]
- Yao J, Chen C, Guo Y, Yang Y, Liu X, Chu S. A review of research on the association between neuron-astrocyte signaling processes and depressive symptoms. Int J Mol Sci 2023; 24(8):6985. doi: 10.3390/ijms24086985 [Crossref] [ Google Scholar]
- Linnerbauer M, Wheeler MA, Quintana FJ. Astrocyte crosstalk in CNS inflammation. Neuron 2020; 108(4):608-22. doi: 10.1016/j.neuron.2020.08.012 [Crossref] [ Google Scholar]
- Li D, Wang Y, Jin X, Hu D, Xia C, Xu H. NK cell-derived exosomes carry miR-207 and alleviate depression-like symptoms in mice. J Neuroinflammation 2020; 17(1):126. doi: 10.1186/s12974-020-01787-4 [Crossref] [ Google Scholar]
- Ji C, Tang Y, Zhang Y, Li C, Liang H, Ding L. Microglial glutaminase 1 deficiency mitigates neuroinflammation associated depression. Brain BehavImmun 2022; 99:231-45. doi: 10.1016/j.bbi.2021.10.009 [Crossref] [ Google Scholar]
- Zhu LL, Li LD, Lin XY, Hu J, Wang C, Wang YJ. Plasma-derived small extracellular vesicles miR-182-5p is a potential biomarker for diagnosing major depressive disorder. Mol Neurobiol 2025; 62(9):11099-111. doi: 10.1007/s12035-025-04948-9 [Crossref] [ Google Scholar]
- Costa-Ferro ZSM, Cunha RS, Rossi EA, Loiola EC, Cipriano BP, Figueiredo JC. Extracellular vesicles derived from mesenchymal stem cells alleviate depressive-like behavior in a rat model of chronic stress. Life Sci 2025; 366-367:123479. doi: 10.1016/j.lfs.2025.123479 [Crossref] [ Google Scholar]
- Liang T, Wu Z, Li J, Wu S, Shi W, Wang L. The emerging double-edged sword role of exosomes in Alzheimer’s disease. Front Aging Neurosci 2023; 15:1209115. doi: 10.3389/fnagi.2023.1209115 [Crossref] [ Google Scholar]
- Ausó E, Gómez-Vicente V, Esquiva G. Biomarkers for Alzheimer’s disease early diagnosis. J Pers Med 2020; 10(3):114. doi: 10.3390/jpm10030114 [Crossref] [ Google Scholar]
- Si ZZ, Zou CJ, Mei X, Li XF, Luo H, Shen Y. Targeting neuroinflammation in Alzheimer’s disease: from mechanisms to clinical applications. Neural Regen Res 2023; 18(4):708-15. doi: 10.4103/1673-5374.353484 [Crossref] [ Google Scholar]
- Rostagno AA. Pathogenesis of Alzheimer’s disease. Int J Mol Sci 2022; 24(1):107. doi: 10.3390/ijms24010107 [Crossref] [ Google Scholar]
- Dubois B, Hampel H, Feldman HH, Scheltens P, Aisen P, Andrieu S. Preclinical Alzheimer’s disease: definition, natural history, and diagnostic criteria. Alzheimers Dement 2016; 12(3):292-323. doi: 10.1016/j.jalz.2016.02.002 [Crossref] [ Google Scholar]
- d’Errico P, Meyer-Luehmann M. Mechanisms of pathogenic tau and Aβ protein spreading in Alzheimer’s disease. Front Aging Neurosci 2020; 12:265. doi: 10.3389/fnagi.2020.00265 [Crossref] [ Google Scholar]
- Liu WL, Lin HW, Lin MR, Yu Y, Liu HH, Dai YL. Emerging blood exosome-based biomarkers for preclinical and clinical Alzheimer’s disease: a meta-analysis and systematic review. Neural Regen Res 2022; 17(11):2381-90. doi: 10.4103/1673-5374.335832 [Crossref] [ Google Scholar]
- Bouwman FH, Frisoni GB, Johnson SC, Chen X, Engelborghs S, Ikeuchi T. Clinical application of CSF biomarkers for Alzheimer’s disease: from rationale to ratios. Alzheimers Dement (Amst) 2022; 14(1):e12314. doi: 10.1002/dad2.12314 [Crossref] [ Google Scholar]
- Eitan E, Hutchison ER, Marosi K, Comotto J, Mustapic M, Nigam SM. Extracellular vesicle-associated Aβ mediates trans-neuronal bioenergetic and Ca2 + -handling deficits in Alzheimer’s disease models. NPJ Aging Mech Dis 2016; 2:16019. doi: 10.1038/npjamd.2016.19 [Crossref] [ Google Scholar]
- Kim KY, Shin KY, Chang KA. Brain-derived exosomal proteins as effective biomarkers for Alzheimer’s disease: a systematic review and meta-analysis. Biomolecules 2021; 11(7):980. doi: 10.3390/biom11070980 [Crossref] [ Google Scholar]
- Kumar A, Su Y, Sharma M, Singh S, Kim S, Peavey JJ. MicroRNA expression in extracellular vesicles as a novel blood-based biomarker for Alzheimer’s disease. Alzheimers Dement 2023; 19(11):4952-66. doi: 10.1002/alz.13055 [Crossref] [ Google Scholar]
- Wang X, Yang H, Liu C, Liu K. A new diagnostic tool for brain disorders: extracellular vesicles derived from neuron, astrocyte, and oligodendrocyte. Front Mol Neurosci 2023; 16:1194210. doi: 10.3389/fnmol.2023.1194210 [Crossref] [ Google Scholar]
- Zou Y, Mu D, Ma X, Wang D, Zhong J, Gao J. Review on the roles of specific cell-derived exosomes in Alzheimer’s disease. Front Neurosci 2022; 16:936760. doi: 10.3389/fnins.2022.936760 [Crossref] [ Google Scholar]
- Koinuma S, Shimozawa N, Yasutomi Y, Kimura N. Aging induces abnormal accumulation of Aβ in extracellular vesicle and/or intraluminal membrane vesicle-rich fractions in nonhuman primate brain. Neurobiol Aging 2021; 106:268-81. doi: 10.1016/j.neurobiolaging.2021.06.022 [Crossref] [ Google Scholar]
- Chen YA, Lu CH, Ke CC, Chiu SJ, Jeng FS, Chang CW. Mesenchymal stem cell-derived exosomes ameliorate Alzheimer’s disease pathology and improve cognitive deficits. Biomedicines 2021; 9(6):594. doi: 10.3390/biomedicines9060594 [Crossref] [ Google Scholar]
- Liu S, Fan M, Xu JX, Yang LJ, Qi CC, Xia QR. Exosomes derived from bone-marrow mesenchymal stem cells alleviate cognitive decline in AD-like mice by improving BDNF-related neuropathology. J Neuroinflammation 2022; 19(1):35. doi: 10.1186/s12974-022-02393-2 [Crossref] [ Google Scholar]
- Liu H, Jin M, Ji M, Zhang W, Liu A, Wang T. Hypoxic pretreatment of adipose-derived stem cell exosomes improved cognition by delivery of circ-Epc1 and shifting microglial M1/M2 polarization in an Alzheimer’s disease mice model. Aging (Albany NY) 2022; 14(7):3070-83. doi: 10.18632/aging.203989 [Crossref] [ Google Scholar]
- Li N, Shu J, Yang X, Wei W, Yan A. Exosomes derived from M2 microglia cells attenuates neuronal impairment and mitochondrial dysfunction in Alzheimer’s disease through the PINK1/Parkin pathway. Front Cell Neurosci 2022; 16:874102. doi: 10.3389/fncel.2022.874102 [Crossref] [ Google Scholar]
- Iyaswamy A, Thakur A, Guan XJ, Krishnamoorthi S, Fung TY, Lu K. Fe65-engineered neuronal exosomes encapsulating corynoxine-B ameliorate cognition and pathology of Alzheimer’s disease. Signal Transduct Target Ther 2023; 8(1):404. doi: 10.1038/s41392-023-01657-4 [Crossref] [ Google Scholar]
- Caire MJ, Reddy V, Varacallo MA. Physiology, synapse. In: StatPearls [Internet]. Treasure Island, FL: StatPearls Publishing; 2024.
- Wang X, Christian KM, Song H, Ming GL. Synaptic dysfunction in complex psychiatric disorders: from genetics to mechanisms. Genome Med 2018; 10(1):9. doi: 10.1186/s13073-018-0518-5 [Crossref] [ Google Scholar]
- Tavakolizadeh J, Roshanaei K, Salmaninejad A, Yari R, Nahand JS, Khoshdel Sarkarizi H. MicroRNAs and exosomes in depression: potential diagnostic biomarkers. J Cell Biochem 2018; 119(5):3783-97. doi: 10.1002/jcb.26599 [Crossref] [ Google Scholar]
- Duarte-Silva E, Oriá AC, Mendonça IP, de Melo MG, Paiva IH, Maes M. Tiny in size, big in impact: extracellular vesicles as modulators of mood, anxiety and neurodevelopmental disorders. NeurosciBiobehav Rev 2022; 135:104582. doi: 10.1016/j.neubiorev.2022.104582 [Crossref] [ Google Scholar]
- Goetzl EJ, Abner EL, Jicha GA, Kapogiannis D, Schwartz JB. Declining levels of functionally specialized synaptic proteins in plasma neuronal exosomes with progression of Alzheimer’s disease. FASEB J 2018; 32(2):888-93. doi: 10.1096/fj.201700731R [Crossref] [ Google Scholar]
- Cui GH, Wu J, Mou FF, Xie WH, Wang FB, Wang QL. Exosomes derived from hypoxia-preconditioned mesenchymal stromal cells ameliorate cognitive decline by rescuing synaptic dysfunction and regulating inflammatory responses in APP/PS1 mice. FASEB J 2018; 32(2):654-68. doi: 10.1096/fj.201700600R [Crossref] [ Google Scholar]
- Christensen DL, Maenner MJ, Bilder D, Constantino JN, Daniels J, Durkin MS. Prevalence and characteristics of autism spectrum disorder among children aged 4 years - early autism and developmental disabilities monitoring network, seven sites, United States, 2010, 2012, and 2014. MMWR SurveillSumm 2019; 68(2):1-19. doi: 10.15585/mmwr.ss6802a1 [Crossref] [ Google Scholar]
- Underwood JF, Del Pozo-Banos M, Frizzati A, Rai D, John A, Hall J. Neurological and psychiatric disorders among autistic adults: a population healthcare record study. Psychol Med 2023; 53(12):5663-73. doi: 10.1017/s0033291722002884 [Crossref] [ Google Scholar]
- Sheldrick RC, Maye MP, Carter AS. Age at first identification of autism spectrum disorder: an analysis of two US surveys. J Am Acad Child Adolesc Psychiatry 2017; 56(4):313-20. doi: 10.1016/j.jaac.2017.01.012 [Crossref] [ Google Scholar]
- Hodges H, Fealko C, Soares N. Autism spectrum disorder: definition, epidemiology, causes, and clinical evaluation. TranslPediatr 2020; 9(Suppl 1):S55-65. doi: 10.21037/tp.2019.09.09 [Crossref] [ Google Scholar]
- Critchfield JW, van Hemert S, Ash M, Mulder L, Ashwood P. The potential role of probiotics in the management of childhood autism spectrum disorders. Gastroenterol Res Pract 2011; 2011:161358. doi: 10.1155/2011/161358 [Crossref] [ Google Scholar]
- Herman S, Fishel I, Offen D. Intranasal delivery of mesenchymal stem cells-derived extracellular vesicles for the treatment of neurological diseases. Stem Cells 2021; 39(12):1589-600. doi: 10.1002/stem.3456 [Crossref] [ Google Scholar]
- Qin Q, Li M, Fan L, Zeng X, Zheng D, Wang H. RVG engineered extracellular vesicles-transmitted miR-137 improves autism by modulating glucose metabolism and neuroinflammation. Mol Psychiatry 2025; 30(9):4072-84. doi: 10.1038/s41380-025-02988-0 [Crossref] [ Google Scholar]
- Guo H, Huang B, Wang Y, Zhang Y, Ma Q, Ren Y. Bone marrow mesenchymal stem cells-derived exosomes improve injury of hippocampal neurons in rats with depression by upregulating microRNA-26a expression. Int Immunopharmacol 2020; 82:106285. doi: 10.1016/j.intimp.2020.106285 [Crossref] [ Google Scholar]
- Guo H, Huang B, Wang Y, Zhang Y, Ma Q, Ren Y. Bone marrow mesenchymal stem cells-derived exosomes improve injury of hippocampal neurons in rats with depression by upregulating microRNA-26a expression. Int Immunopharmacol 2020; 82:106285. doi: 10.1016/j.intimp.2020.106285 [Crossref] [ Google Scholar]
- Liang Y, Duan L, Xu X, Li X, Liu M, Chen H. Mesenchymal stem cell-derived exosomes for treatment of autism spectrum disorder. ACS Appl Bio Mater 2020; 3(9):6384-93. doi: 10.1021/acsabm.0c00831 [Crossref] [ Google Scholar]
- Logue MW, van Rooij SJ, Dennis EL, Davis SL, Hayes JP, Stevens JS. Smaller hippocampal volume in posttraumatic stress disorder: a multisite ENIGMA-PGC study: subcortical volumetry results from posttraumatic stress disorder consortia. Biol Psychiatry 2018; 83(3):244-53. doi: 10.1016/j.biopsych.2017.09.006 [Crossref] [ Google Scholar]
- Ousdal OT, Milde AM, Hafstad GS, Hodneland E, Dyb G, Craven AR. The association of PTSD symptom severity with amygdala nuclei volumes in traumatized youths. Transl Psychiatry 2020; 10(1):288. doi: 10.1038/s41398-020-00974-4 [Crossref] [ Google Scholar]
- Kang HJ, Yoon S, Lee S, Choi K, Seol S, Park S. FKBP5-associated miRNA signature as a putative biomarker for PTSD in recently traumatized individuals. Sci Rep 2020; 10(1):3353. doi: 10.1038/s41598-020-60334-6 [Crossref] [ Google Scholar]
- Lee MY, Baxter D, Scherler K, Kim TK, Wu X, Abu-Amara D. Distinct profiles of cell-free microRNAs in plasma of veterans with post-traumatic stress disorder. J Clin Med 2019; 8(7):963. doi: 10.3390/jcm8070963 [Crossref] [ Google Scholar]
- Bam M, Yang X, Busbee BP, Aiello AE, Uddin M, Ginsberg JP. Increased H3K4me3 methylation and decreased miR-7113-5p expression lead to enhanced Wnt/β-catenin signaling in immune cells from PTSD patients leading to inflammatory phenotype. Mol Med 2020; 26(1):110. doi: 10.1186/s10020-020-00238-3 [Crossref] [ Google Scholar]
- Guo J, Chen S, Li F, Hou S, Guo M, Yuan X. CXCL8 delivered by plasma-derived exosomes induces the symptoms of post-traumatic stress disorder through facilitating astrocyte-neuron communication. J Psychiatr Res 2023; 161:261-72. doi: 10.1016/j.jpsychires.2023.02.040 [Crossref] [ Google Scholar]
- Mokhothu TM, Tanaka KZ. Characterizing hippocampal oscillatory signatures underlying seizures in temporal lobe epilepsy. Front BehavNeurosci 2021; 15:785328. doi: 10.3389/fnbeh.2021.785328 [Crossref] [ Google Scholar]
- Bandopadhyay R, Singh T, Ghoneim MM, Alshehri S, Angelopoulou E, Paudel YN. Recent developments in diagnosis of epilepsy: scope of microRNA and technological advancements. Biology (Basel) 2021; 10(11):1097. doi: 10.3390/biology10111097 [Crossref] [ Google Scholar]
- Batool A, Hill TD, Nguyen NT, Langa E, Diviney M, Mooney C. Altered biogenesis and microRNA content of hippocampal exosomes following experimental status epilepticus. Front Neurosci 2019; 13:1404. doi: 10.3389/fnins.2019.01404 [Crossref] [ Google Scholar]
- Gitaí DL, Dos Santos YD, Upadhya R, Kodali M, Madhu LN, Shetty AK. Extracellular vesicles in the forebrain display reduced miR-346 and miR-331-3p in a rat model of chronic temporal lobe epilepsy. Mol Neurobiol 2020; 57(3):1674-87. doi: 10.1007/s12035-019-01797-1 [Crossref] [ Google Scholar]
- Lin Z, Gu Y, Zhou R, Wang M, Guo Y, Chen Y. Serum exosomal proteins F9 and TSP-1 as potential diagnostic biomarkers for newly diagnosed epilepsy. Front Neurosci 2020; 14:737. doi: 10.3389/fnins.2020.00737 [Crossref] [ Google Scholar]
- Yu T, Fu H, Sun JJ, Ding DR, Wang H. miR-106b-5p upregulation is associated with microglial activation and inflammation in the mouse hippocampus following status epilepticus. Exp Brain Res 2021; 239(11):3315-25. doi: 10.1007/s00221-021-06208-3 [Crossref] [ Google Scholar]
- Zhang W, Wang T, Xue Y, Zhan B, Lai Z, Huang W. Research progress of extracellular vesicles and exosomes derived from mesenchymal stem cells in the treatment of oxidative stress-related diseases. Front Immunol 2023; 14:1238789. doi: 10.3389/fimmu.2023.1238789 [Crossref] [ Google Scholar]
- Abhijna Ballal R, Shivakumar Reddy K, Chandran D, Hegde S, Upadhya R, Se PK. Cell-specific extracellular vesicle-encapsulated exogenous GABA controls seizures in epilepsy. Stem Cell Res Ther 2024; 15(1):108. doi: 10.1186/s13287-024-03721-4 [Crossref] [ Google Scholar]
- Tondo L, Vázquez GH, Baldessarini RJ. Depression and mania in bipolar disorder. CurrNeuropharmacol 2017; 15(3):353-8. doi: 10.2174/1570159x14666160606210811 [Crossref] [ Google Scholar]
- Ceylan D, Tufekci KU, Keskinoglu P, Genc S, Özerdem A. Circulating exosomal microRNAs in bipolar disorder. J Affect Disord 2020; 262:99-107. doi: 10.1016/j.jad.2019.10.038 [Crossref] [ Google Scholar]
- Delwing-de Lima D, Cyrino LAR, Ferreira GK, Delwing-Dal Magro D, Calegari CR, Cabral H. Neuroinflammation and neuroprogression produced by oxidative stress in euthymic bipolar patients with different onset disease times. Sci Rep 2022; 12(1):16742. doi: 10.1038/s41598-022-21170-y [Crossref] [ Google Scholar]
- Fries GR, Lima CN, Valvassori SS, Zunta-Soares G, Soares JC, Quevedo J. Preliminary investigation of peripheral extracellular vesicles’ microRNAs in bipolar disorder. J Affect Disord 2019; 255:10-4. doi: 10.1016/j.jad.2019.05.020 [Crossref] [ Google Scholar]
- Thakur A, Rai D. Global requirements for manufacturing and validation of clinical grade extracellular vesicles. J Liq Biopsy 2024; 6:100278. doi: 10.1016/j.jlb.2024.100278 [Crossref] [ Google Scholar]
- Wiest EF, Zubair AC. Generation of current good manufacturing practices-grade mesenchymal stromal cell-derived extracellular vesicles using automated bioreactors. Biology (Basel) 2025; 14(3):313. doi: 10.3390/biology14030313 [Crossref] [ Google Scholar]